Zellatmung: Funktion, Was ist Zellatmung, wofür ist es wichtig
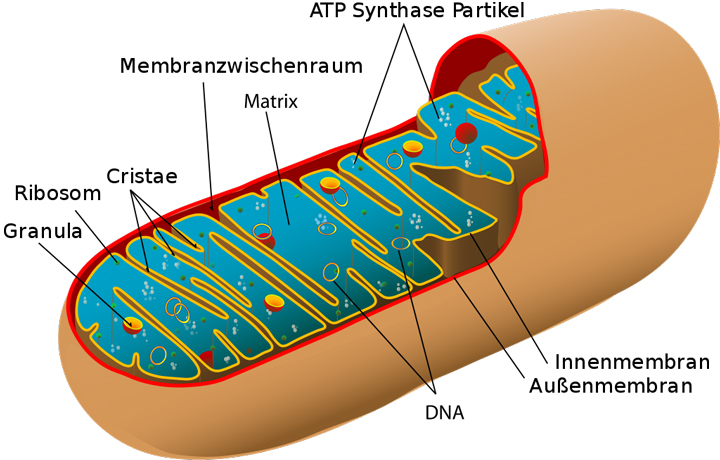
Mitochondrien sind die Orte der Zellatmung, Abbild eines tierischen Mitochondriums mit Innen- und Außenmembran
Zellatmung ist ein Stoffwechselvorgang, um Energie zu gewinnen. Etwas genauer gesagt: Unter dem Begriff der Zellatmung versteht man die Bildung von Adenosintriphosphat (ATP) unter Verbrauch von Betriebsstoffen wie z.B. Glucose und molekularem Sauerstoff. Dieser Prozess findet in den Mitochondrien der Zellen statt.
Inhalt
Mitochondrien – Der Ort der Zellatmung
Die Zellatmung findet laut Definition in den Mitochondrien statt. Dennoch beginnt der Abbau von Betriebsstoffen wie Glucose bereits im Cytosol (flüssiger Raum zwischen Zellorganellen) statt. Die aus diesen ersten Schritten des Abbaus entstandenen, energiereichen Moleküle, gelangen daraufhin in die Mitochondrien.
Mitochondrien sind kleine Zellorganellen, mit einem Durchmesser von ca. 1,5 µm und einer Länge von etwa 2-8 µm. Ihre Größe gleicht somit einer typischen Bakterienzelle. Eine weitere Eigenschaft ist ihre Fähigkeit, sich unabhängig vom Zellkern teilen zu können. Innerhalb von Zellen ist die Anzahl an Mitochondrien sehr variabel. In Eizellen können mehrere hunderttausend Mitochondrien vorkommen, während in einigen einzelligen Organismen nur ein einziges Mitochondrium vorkommt. Grundsätzlich gilt: Je mehr Energie eine Zelle benötigt, desto mehr Mitochondrien enthält sie auch.
Strukturell bestehen Mitochondrien aus zwei Membranen. Die äußere Membran ist glatt und weist eine Schutzfunktion auf. Im Inneren ist eine zweite Membran angeheftet, die durch viele Einstülpungen eine deutlich größere Oberfläche aufweist als die äußere Membran. Durch die Regelmäßigkeit der Einstülpungen wurden diese zu Strukturen zusammengefasst, die man als Cristae bezeichnet. Die innere Membran kontrolliert darüber hinaus verstärkt das Einströmen von Substanzen und ihr innewohnend sind viele Proteinkomplexe, die an der Zellatmung beteiligt sind. Durch die größere Oberfläche der Cristae entsteht somit eine erhöhte Effizienz der Zellatmung, da mehr Proteinkomplexe eingebettet werden können als in einer glatten Membran.
Der Raum, der von der inneren Membran begrenzt wird, wird als Mitochondrienmatrix bezeichnet. Sie beinhaltet eigene Ribosomen (Organellen, die an der Proteinsynthese beteiligt sind) und DNA. Somit sind Mitochondrien in der Lage, eigene Proteine zu synthetisieren, um die Zellatmung aufrechterhalten zu können. Die DNA der Mitochondrien ist ein Rest eines größeren Chromosoms eines Prokaryoten (Zelle ohne Zellkern, z.B. Bakterien), der im Laufe der Evolution von einer eukaryotischen Zelle (Zelle mit Zellkern) aufgenommen wurde. Dies würde auch die Größe der Mitochondrien erklären.
Was bedeutet Zellatmung
Zellatmung bedeutet, dass Betriebsstoffe innerhalb von Zellen unter Sauerstoffverbrauch abgebaut werden, um ATP zu generieren. Dies geschieht über Redoxreaktionen. Redoxreaktionen sind chemische Reaktionen, bei denen ein Reaktionspartner reduziert wird (Elektronen aufnimmt) und ein anderer Reaktionspartner oxidiert wird (Elektronen abgibt). Im Umkehrschluss sind Oxidationsmittel jene Reaktionspartner, die während einer Redoxreaktion reduziert werden und Reduktionsmittel jene, die oxidiert werden. Bei der Zellatmung fungiert Glucose als Reduktionsmittel und Sauerstoff als Oxidationsmittel.
Die Zellatmung beinhaltet mehrere Stoffwechselwege. Dazu zählen die Glykolyse, die Pyruvatoxidation, der Citratzyklus und die Atmungskette. Nach Beendigung aller Stoffwechselwege wird die anfänglich zur Verfügung stehende Glucose vollständig zu CO2 und Wasser oxidiert. Neben der aeroben (Vorhandensein von Sauerstoff) Zellatmung sind Zellen darüber hinaus dazu befähigt, unter anaeroben (Fehlen von Sauerstoff) Bedingungen Energie zu gewinnen. Die anaeroben Prozesse werden hier jedoch nicht weiter erläutert.
Wo und wann findet Zellatmung statt
Prinzipiell findet die Zellatmung bei Eukaryoten zu jeder Zeit im Cytosol und in den Mitochondrien aller Zellen statt, da die daraus gewonnene Energie, die in ATP gespeichert wird, bei vielen anderen ablaufenden Prozessen innerhalb der Zelle von Nöten ist. Dennoch kann man die genaue Lokalisation der ablaufenden Prozesse exakter bestimmen. Bei Eukaryoten findet die Glykolyse und die anaerobe Gärung im Cytosol statt, da beide Prozesse unter anaeroben Bedingungen stattfinden. Die Pyruvatoxidation und die Atmungskette findet bei Eukaryoten an der inneren Mitochondrienmembran statt, der Citratzyklus jedoch in der Mitochondrienmatrix.
Bei Prokaryoten laufen die Prozesse nur an zwei Orten ab. Im Cytoplasma (der flüssige Raum innerhalb einer Zelle) finden die Glykolyse, die Gärung und der Citratzyklus statt. Die Pyruvatoxidation und die Atmungskette laufen an der Plasmamembran ab. Auch bei Prokaryoten findet die Zellatmung grundsätzlich immer dann statt, wenn Energie benötigt wird. Bei begeißelten Bakterien kann dies beispielsweise der Fall sein, wenn sich die Bakterienzelle durch den Geißelschlag fortbewegt.
Die genauen Stoffwechselwege der Zellatmung
Glykolyse
Der erste Schritt der Zellatmung ist die Glykolyse. Während der Glykolyse wird aus Glucose (ein Kohlenhydrat, C6-Körper) in Pyruvat (auch bekannt als Salz der Brenztraubensäure, C3-Körper) gewonnen. Hierbei spielen mehrere Reaktionen eine wichtige Rolle. Diese kann man in eine ATP-Investitionsphase und eine ATP-Gewinnphase gliedern.
ATP-Investitionsphase der Glykolyse
Die Glucose wird zuerst durch das Enzym Hexokinase phosphoryliert (ein Phosphatrest wird angehängt). Hierbei wird ein ATP-Molekül verbraucht und in ADP umgewandelt, da der übertragene Phosphatrest vom ATP stammt.
Hierbei entsteht Glucose-6-Phosphat, welches durch das Enzym Phosphoglucoisomerase in Fructose-6-Phosphat umstrukturiert wird.
Daraufhin wird durch das Enzym Phosphofructokinase ein Molekül Fructose-1,6-Bisphosphat phosphoryliert. Hierbei wird wieder ein Phosphatrest von ATP übertragen, das zu ADP reagiert.
Fructose liegt meist in einer Ringstruktur vor, welche eine hohe energetische Stabilität aufweist. Diese muss im nächsten Schritt geöffnet werden, um die Weiterverarbeitung zu gewährleisten. Diese Reaktion wird durch das Enzym Aldolase katalysiert. Aus der Öffnung des Fructoserings entstehen jeweils ein Molekül Dihydroxyacetonphosphat (DHAP) und ein Molekül Glycerinaldehyd-3-Phosphat (G3P).
Weiterhin finden während der Glykolyse jedoch nur Reaktionen mit G3P statt, weshalb aus DHAP ein Molekül G3P entstehen muss. Diese Reaktion katalysiert das Enzym Isomerase. Als Zwischenbilanz der ATP-Investitionsphase wurden aus einem Molekül Glucose zwei Moleküle G3P, wobei insgesamt pro Molekül Glucose jeweils 2 ATP verbraucht wurden. Daraufhin folgt die ATP-Gewinnungsphase.
ATP-Gewinnungsphase der Glykolyse
Die beiden, während der ATP-Investitionsphase entstandenen, G3P Moleküle werden mithilfe des Enzyms Triosephosphatdehydrogenase oxidiert und nehmen gleichzeitig einen Phosphatrest auf. Während der Oxidation wird ein Wasserstoffatom abgespalten, welches ein Molekül NAD+ (Nikotinamidadenindinukleotid) zu NADH reduziert. Aus dieser Reaktion resultieren zwei Moleküle 1,3-Bisphosphoglycerat (BPG).
Im nächsten Schritt übertragen die beiden BPG-Moleküle mithilfe des Enzyms Phosphoglyceratkinase jeweils eine Phosphatgruppe am C1-Atom auf ein Molekül ADP (Adenosindiphosphat). Hierdurch entstehen zwei Moleküle ATP.
Durch die Abspaltung des Phosphatrestes im vorangegangenen Reaktionsschritt entstehen zwei Moleküle 3-Phosphoglycerat (3PG). Diese werden durch das Enzym Phosphoglyceromutase zu zwei Molekülen 2-Phosphoglycerat (2PG) mutiert, indem der Phosphatrest umgelagert wird.
Im nächsten Reaktionsschritt wird mithilfe des Enzyms Enolase pro Molekül 2PG ein Molekül Wasser abgespalten, wodurch zwei Moleküle Phosphoenolpyruvat (PEP) entstehen.
Das Enzym Pyrvatkinase überträgt im nächsten Schritt den Phosphatrest der beiden Moleküle PEP auf zwei Moleküle ADP, wodurch 2 ATP entstehen. Dadurch entsteht das finale Produkt der Glykolyse, das Pyruvat, in doppelter Ausführung.
Die ATP-Bilanz beträgt nach der Glykolyse insgesamt +2, da zunächst zwei ATP investiert wurden, in darauffolgenden Schritten jedoch 4 ATP generiert wurden.
Pyruvatoxidation
Die Pyruvatoxidation ist für den darauffolgenden Citratzyklus essenziell. Hierbei werden die beiden Pyruvatmoleküle aus der Glykolyse in zwei Moleküle Acetat oxidiert. Daraufhin werden die beiden Moleküle Acetat zu Acetyl-CoA (CoA = Coenzym A) umgelagert, das eine Schlüsselrolle im Citratzyklus spielt.
Citratzyklus
Der Citratzyklus beginnt mit der Reaktion von einem Molekül Acetyl-CoA und einem Molekül Oxalacetat. Nach Abspaltung des CoA reagieren beide Moleküle zusammen zu Citrat. Das abgespaltene CoA kann von dem Enzym Pyruvatdehydrogenase weiter verwendet werden, um Pyruvat zu decarboxylieren (ein Molekül CO2 abspalten).
Citrat wird im Laufe der folgenden Reaktion zu Isocitrat umstrukturiert.
In der nächsten Reaktion werden sowohl ein Molekül NAD+ zu NADH reduziert als auch Isocitrat zu alpha-Ketoglutarat oxidiert und darüber hinaus wird ein Molekül CO2 abgespalten.
Während dem vierten Reaktionsschritt wird das alpha-Ketoglutarat mithilfe des CoA zu Succinyl-CoA oxidiert. Gleichzeitig wird ein Molekül NAD+ zu NADH reduziert und erneut ein Molekül CO2 abgespalten.
Das Succinyl-CoA wird im nächsten Reaktionsschritt das CoA abspalten und wird zu Succinat (Salz der Bernsteinsäure). Durch die Abspaltung des CoA wird Energie frei, die dazu genutzt wird zunächst GDP (Guanosindiphosphat) in GTP (Guanosintriphosphat) umzuwandeln. Das GTP überträgt unmittelbar danach den dritten Phosphatrest auf ein Molekül ADP, wodurch ein Molekül ATP entsteht.
Aus dem Succinat wird mittels Oxidation Fumarat (Salz der Fumarsäure). Dabei wird ein Molekül FAD zu FADH2 reduziert.
Im nächsten Schritt findet eine Reaktion von Wasser und Fumarat statt, die die Entstehung von Malat zur Folge hat. Das entstandene Malat reagiert durch Oxidation zu Oxalacetat. Hierbei wird erneut ein Molekül NAD+ zu NADH reduziert. Da im letzten Schritt das ursprünglich verwendete Oxalacetat regeneriert wurde, kann der Citratzyklus erneut durchlaufen werden.
Insgesamt wird der Citratzyklus pro Glucosemolekül, das die Glykolyse durchläuft, zwei Mal durchlaufen (einmal pro Pyruvatmolekül, das entsteht).
Atmungskette
Die Atmungskette ist der letzte Schritt der Zellatmung, wobei weitere ATP-Moleküle generiert werden. Während der weiteren Synthese (Herstellung) von ATP werden sog. Elektronencarrier reoxidiert, weshalb man diesen Prozess auch als oxidative Phosphorylierung bezeichnet. Grundsätzlich lässt sich der Prozess der oxidativen Phosphorylierung in zwei Teilschritte gliedern: Den Elektronentransport und die Chemiosmose.
Während des Elektronentransports fließen Elektronen, die vom NADH und FADH2 stammen, über membranassoziierte (in der Membran liegende) Elektronentransporter. Diese erzeugen einen Protonenausstrom aus der Mitochondrienmatrix, wodurch eine Protonengradient (Ungleichgewicht) entsteht.
Bei der Chemiosmose wird der Protonengradient dazu genutzt, dass die Protonen durch einen gewissen Kanal, die ATP-Synthase (Enzym zur Herstellung von ATP), zurück in die Mitochondrienmatrix diffundieren. Die ATP-Synthase ist die einzige Möglichkeit für die Protonen, wieder in die Mitochondrienmatrix zurückzugelangen, da diese sonst für Protonen undurchlässig ist.
Die Atmungskette ist ein sehr komplexer Prozess, weshalb man sich die Frage stellen könnte, wieso innerhalb der Zelle nicht direkt NADH mit Sauerstoff zu NAD+ und Wasser reagiert. Bei dieser Reaktion würde so viel Energie frei, dass es einer Explosion innerhalb der Zelle gleichkäme. Es gibt auch keinen bekannten Stoffwechselweg, der den größten Teil der dort freiwerdenden Energie absorbieren (aufnehmen) würde. Somit bleibt der Zelle nur die Atmungskette als letzte Möglichkeit.
Wie im Abschnitt „Wo und wann findet Zellatmung statt“ erwähnt, findet die Atmungskette bei Eukaryoten in der inneren Mitochondrienmembran statt, bei Prokaryoten in der Plasmamembran.
Um den Prozess der Atmungskette noch genauer verstehen zu können, sollte man sich einen detaillierteren Überblick über die beteiligten Proteinkomplexe und andere Moleküle verschaffen. Während des Elektronentransports werden zunächst vier große Proteinkomplexe mit ihren assoziierten Enzymen durchlaufen. Zwischen den vier großen Proteinkomplexen gibt es noch zwei weitere Moleküle, das Cytochrom c und Ubichinon (wird meist mit Q abgekürzt).
Der Prozess der Atmungskette startet damit, dass NADH Elektronen über den Proteinkomplex 1 (NADH-Q-Oxidoreduktase) an Ubichinon weitergibt. Hierbei wird eine beachtliche Menge Energie frei.
Komplex 2 (Succinatdehydrogenase) übernimmt Elektronen des FADH2, das aus dem Citratzyklus entspringt, und überträgt die Elektronen auch an Ubichinon. Diese Elektronen werden jedoch deutlich später in die Atmungskette eingeschleust und erzeugen im Endeffekt weniger ATP als die Elektronen des NADH. Jedes Elektronenpaar, das von NADH auf Sauerstoff übertragen wird, generiert ca. 2,5 ATP, wohingegen jedes Elektronenpaar aus FADH2 nur 1,5 ATP erzeugt.
Der dritte Komplex (Cytochrom-c-Oxidoreduktase) erhält nun die Elektronen vom reduzierten Ubichinon und leitet sie an das Cytochrom c weiter.
Der vierte Komplex (Cytochrom-c-Oxidase) empfängt die Elektronen vom Cytochrom c und überträgt sie auf ein Sauerstoffmolekül, das daraufhin zu Wasser reduziert wird.
Bei diesen Reaktionen werden zwei Protonen verbraucht, was dazu beiträgt, dass ein Protonengradient im inneren entsteht. Darüber hinaus werden auch weitere Protonen aktiv durch die Mitochondrienmembran in den Intermembranraum (Raum zwischen der inneren und äußeren Membran) gepumpt. Dies geschieht durch die Komplexe 1,3 und 4. Der Protonengradient dient somit als Quelle potenzieller Energie, die zur ATP-Synthese genutzt wird.
Weiterhin erzeugt der Protonengradient aufgrund der positiven Ladung auch eine Änderung der elektrischen Ladungsverteilung. Somit ist die Matrix negativer geladen als der Intermembranraum. Durch diese Ladungsverschiebung steht weitere potenzielle Energie zur ATP-Synthese zur Verfügung.
Die Chemiosmose nutzt die ATP-Synthase zur Verbindung der Protonendiffusion mit der ATP-Synthese. Da die ATP-Synthese eine reversible (rückgängig zu machende) Reaktion ist, kann sie auch in zwei Richtungen ablaufen. Entweder wird ADP unter Nutzung von Energie zu ATP oder ATP wird zu ADP hydrolysier, wodurch Energie frei wird.
Bilanz der Zellatmung inklusive aller Reaktionen
Insgesamt werden 10 NADH+H+, 2 FADH2 , 34 ADP, 34 Phosphatresten und 6 Moleküle Sauerstoff verbraucht, woraus insgesamt 34 ATP, 12 Moleküle Wasser, 10 NAD+ und 2 FAD generiert werden.
Wie betreiben die unterschiedlichen Lebewesen Zellatmung
Unterschiedliche Domänen weisen bei detaillierter Betrachtung teilweise unterschiedliche Stoffwechselwege auf, wobei die grundlegenden Reaktionen ähnlich sind. Dies hängt auch immer davon ab, unter welchen Bedingungen die jeweiligen Organismen leben.
Unter aeroben Bedingungen findet bei allen Lebewesen, die unter aeroben Bedingungen leben können, die normale Zellatmung statt. Für viele obligat anaerobe (Organismen, die hauptsächlich unter niedrigen Sauerstoffkonzentrationen bzw. unter Ausschluss von Sauerstoff leben) Bakterien oder Archaeen ist die Wirkung von Sauerstoff giftig. Nicht direkt die Anwesenheit des Sauerstoffs wirkt giftig, sondern die giftigen Derivate (Stammverbindung mit funktioneller Gruppe) des Sauerstoffs können zellschädigend wirken.
Eine weitere Abwandlung der anaeroben Gärung ist die der Hefe. Die Hefe, als Vertreter der Pilze, ist dazu in der Lage, unter Ausschluss von Sauerstoff Alkohol (hauptsächlich Ethanol) zu erzeugen. Tierische Zellen würden unter anaeroben Bedingungen jedoch aus Pyruvat Milchsäure gären.
Auch an Land lebende Pflanzen betreiben Zellatmung, um Energie aus der, während der Photosynthese entstandenen, Glucose gewinnen zu können. Durch die permanente Verfügbarkeit von Sauerstoff finden bei Pflanzen die gleichen Prozesse statt, wie sie bei allen anderen Organismen unter aeroben Bedingungen stattfinden würden.
Da die ATP-Gewinnung unter aeroben Bedingungen deutlich effizienter abläuft als unter anaeroben Bedingungen, haben viele Tiere eine permanente Sauerstoffversorgung etabliert. Seien es die Kiemen bei Fischen, die Tracheen bei Insekten oder die Lungen bei Amphibien, Vögeln, Reptilien oder Säugetieren. Hierdurch kann die, für die aerobe Zellatmung benötigte, Sauerstoffkonzentration aufrechterhalten werden, um den Körper mit Energie zu versorgen.
Über den Autor
