Aufbau einer Blüte: Bestandteile, Funktionsweise | Pflanzenanatomie
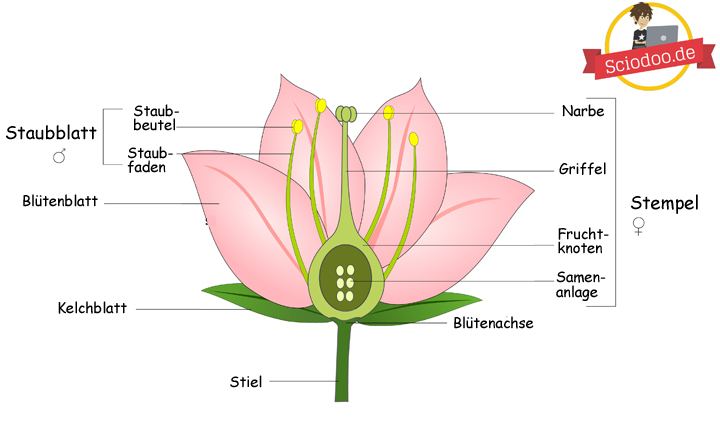
,Grundlegender Aufbau von Blüten einer Blütenpflanze
Der Aufbau von Blüten einer Pflanze ist bereits Thema der Grundschule und wird spätestens in der 5. Klasse vermittelt. In diesen Klassenstufen wird der prinzipielle Aufbau – mit dem Stempel und dem Fruchtblatt – gelehrt. Aber diese Blütenorgane können weiter differenziert werden. Doch zunächst müssen wir abklären, was eine Blüte ist und wofür diese eigentlich zuständig ist.
Inhalt
Was gehört zur Blüte bei einer Blütenpflanze
Um eine Blüte zu definieren, gibt es mehrere Möglichkeiten. Einerseits kann eine Blüte als kurzer Spross angesehen werden, der nur begrenzt wächst und dessen Blätter bei der sexuellen Vermehrung von Pflanzen beteiligt sind. Die indirekte Beteiligung an der Fortpflanzung geschieht durch die Anlockung von Bestäubern, die direkte Beteiligung geschieht durch die Ausbildung von Staub- und Fruchtblättern.

Ein Schmetterling auf der Blüte einer Kamille, angelockt durch Farbe, Geruch und schmackhaften Nektar soll dieser als Bestäuber dienen und so die Fortpflanzung der Kamillenpflanze vorantreiben
Andererseits kann man sich nur auf die Blüten von Angiospermen (Bedecktsamern) beziehen, bei denen eine strikte Gliederung in verschiedene Teilorgane der Blüte erfolgt. Die Angiospermen-Blüte besteht aus einer Blütenhülle, dem Androeceum, dem Gynoeceum und der Blütenachse. Die Blütenhülle wird auch als Perianth bezeichnet und kann entweder einzeln als Perigon oder getrennt in Kelch- und Kronblättern vorliegen.
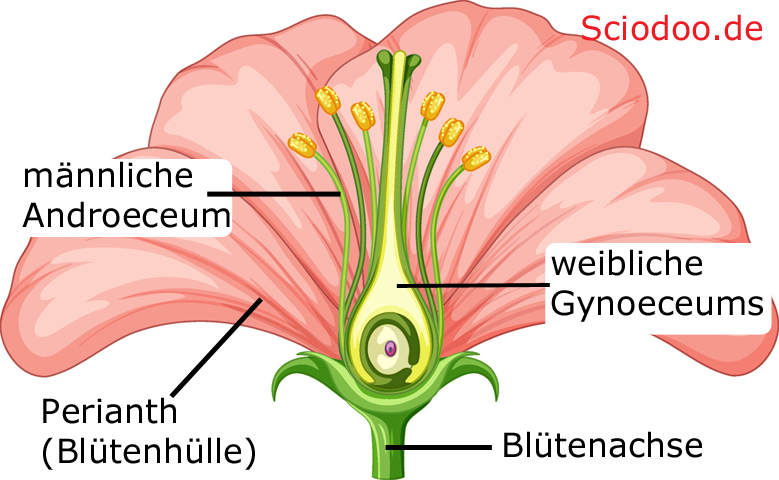
Haupt-Bestandteile einer Blüte mit weiblichen und männlichen Geschlechtsanteilen, einer Blütenhülle zum Schutz und einer Blütenachse als Verbindung zur restlichen Pflanze
Das Androeceum beinhaltet die Staubblätter, das Gynoeceum die Fruchtblätter. Gelangt der Pollen, der sich im Androeceum befindet, per Bestäubung auf die Narbe der Fruchtblätter, so wird die dort befindliche Eizelle befruchtet und ein Embryo entsteht, der dann zum Pflanzensamen heranreift. Im gleichen Zuge wird aus der Blüte eine Frucht. Die Blütenachse bildet den Blütenboden aus.
Aufbau einer Blüte
Der exakte Aufbau einer Blüte lässt sich in die Blütenhülle, die Mikrosporophylle, die Megasporophylle, die Blütenachse und die Nektarien unterteilen. Bei der Bestimmung von Pflanzen spielt allerdings auch die Blattstellung der Blüten eine entscheidende Rolle.
Die Blütenhülle
Die Blütenhülle, die in der Literatur auch als Perianth bezeichnet wird, beinhaltet fortpflanzungsunfähige (sterile) Blätter. Sie existiert nur bei den Gnetopsida und allen Angiospermen. Gnetopsida sind eine Pflanzenklasse, deren Verwandtschaftsverhältnisse noch ungeklärt sind. Dennoch weisen sie Merkmale auf, die darauf schließen lassen, dass sie eher mit den Coniferopyhtina (Abteilung der Gymnospermen) als mit den Angiospermen verwandt sind.

Die Blüte der Tulpe hat keine Differenzierung in Kelch- und Kronen bzw. Blütenblätter, weshalb man diese Blatttyp als Perigonblatt bezeichnet. Die Blütenhülle, also die Gesamtheit der Perigonblätter, wird als Tepalum bezeichnet.
Bei einer gleichförmigen Gestaltung der Blütenhüllblätter wird von einem Perigon gesprochen und die einzelnen Blätter als Tepalen bezeichnet. Hierbei kann das Perigon auch aus mehreren Umläufen von Tepalen bestehen. In diesem Fall wird die Blüte homochlamydeisch genannt.
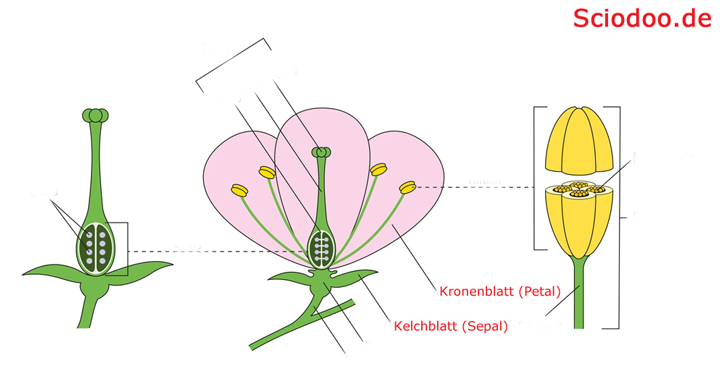
Eine heterochlamydeische Blüte besitzt differenzierte Kelch- und Kronenblätter
Im Falle von heterochlamydeischen Blüten liegt jedoch eine Trennung zwischen Kelch- und Kronenblättern vor. Die äußeren Blätter (Kelchblätter) werden in diesem Fall als Sepalen bezeichnet, die inneren Blätter (Kronblätter) als Petalen. Die Kelchblätter bilden hierbei den Kelch (Calyx), während die Kronblätter die Krone (Corolla) bilden.
Die Staubblätter
Die Staubblätter sind jene Blätter, die die Sporangien (Pollensäcke) tragen. Innerhalb der Sporangien findet die Bildung der männlichen Mikrosporen statt. Die Erscheinung der Staubblätter ist je nach Pflanzengruppe äußerst unterschiedlich. Bei den Nadelbäumen (Koniferen) ist die männliche Blüte zapfenartig gestaltet und besitzt mehrere Staubblätter, die wiederum zwischen zwei bis 20 Pollensäcke beinhalten.
Weiterhin werden die Staubblätter der Angiospermen auch als Stamina bezeichnet. Gegliedert werden die Stamina in einen Staubfaden, auch Filament genannt, und in einen Staubbeutel, der als Anthere bezeichnet wird. Zusammenfassend werden alle Staubblätter als Androeceum bezeichnet.
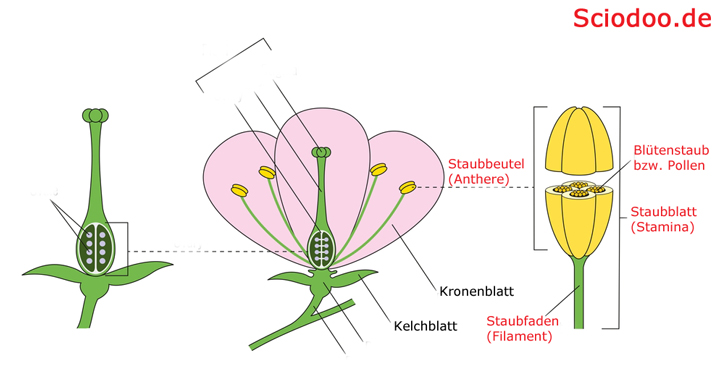
Die Staubblätter sind Teil des männlichen Androeceum und bestehen aus dem Staubbeutel, dem Staubfaden und den Pollen, welche auch als Blütenstaub bezeichnet werden
Die Fruchtblätter
Die Megasporophylle werden nur bei Angiospermen auch als Fruchtblätter bezeichnet. Sie beinhalten die Samenanlagen, aus denen die weiblichen Fortpflanzungsorgane hervorgehen.
Als Gegenbeispiel dienen die Palmfarne, bei denen die Samenanlagen direkt auf Blättern zu finden sind. Hierbei erscheint ein Megasporophyll gestielt und trägt zwei Samenanlagen. Im Vergleich dazu befinden sich die Megasporophylle bei Koniferen auf einer Samenschuppe. Eine Ausnahme bildet hierbei die Eibe, bei der die Samenanlage an einem Kurztrieb steht.
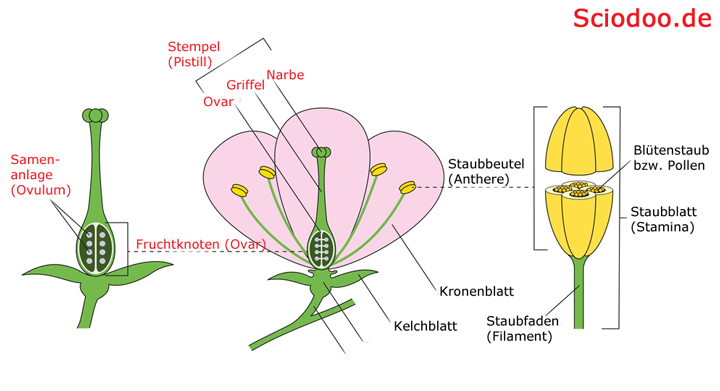
Aufbau eines Fruchtblatts mit Fruchtknoten (Ovar), Samenanlage (Ovulum), Griffel (Stylus), Narbe (Stigma), Stempel (Pistill)
Bei Angiospermen hingegen sind die Samenanlagen in die Fruchtblätter (Karpelle) eingebettet, weshalb sie auch Bedecktsamer genannt werden. Als Gynoeceum bezeichnet man zusammenfassend alle Karpelle einer Blüte. Ein Karpell kann in zwei Bereiche unterteilt werden, in die Stiel- und die Schlauchzone. Innerhalb der Schlauchzone liegen die Samenanlagen.
Ähnlich wie beim Menschen setzen auch bei Pflanzen die Samenanlagen an einer Plazenta an. Über einen sogenannten Griffel ist die Samenanlage mit der Narbe verbunden, an der die Pollen empfangen werden. Weiterhin können Fruchtblätter frei vorliegen, wobei man von einem apokarpen Gynoeceum spricht, aber auch zusammengewachsen vorliegen, was einem coenokarpen Gynoeceum entspricht.
Zeugungsunfähige, sterile Blüten, besitzen hingegen zurückgebildete Staub- und Fruchtblätter, die lediglich eine Anlockfunktion aufweisen.
Die Blütenachse
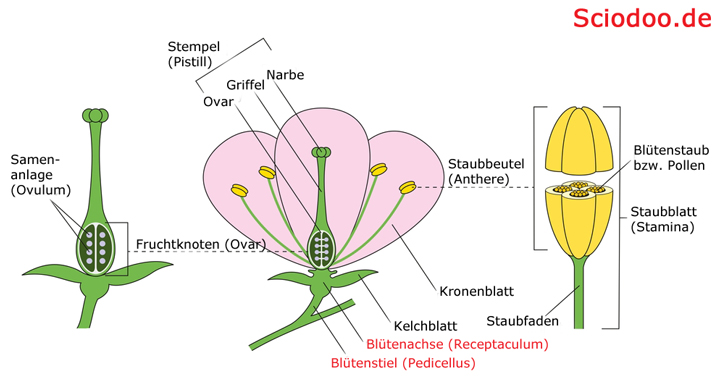
Blütenstiel (Pedicellus) und Blütenachse (Receptaculum) ist die Verbindung zwischen Blütenblättern und der Sprossachse der Pflanze
Die Blütenblätter sind mit der Blütenachse, dem Receptaculum, verbunden. Hierbei können verschiedenste Bereiche der Blüte versenkt oder ausgestreckt werden. Bei Passionsblumen werden z.B. Gynoeceum und Androeceum herausgestreckt. Bei Korbblütlern (Asteraceae), wie z.B. dem Gänseblümchen werden auch ringförmige Strukturen um das Blütenzentrum gestreckt, was zur typischen Blütenform der Korbblütler führt.
Die Nektarien
Um die Bestäubung der Blüten für die Bestäuber attraktiv zu gestalten, bilden Pflanzen Nektarien aus. Diese bestehen aus Drüsengewebe, die Nektar sezernieren. Nektar besteht in der Regel aus Zuckern und Proteinen. Er hat die Hauptaufgabe, Bestäuber anzulocken, die daraufhin unabsichtlich die Blüten bestäuben.
Blütenstellung
Grundsätzlich wird bei der Blütenstellung zwischen zwei Formen, der schraubigen und der wirteligen Form, unterschieden. Die schraubige Anordnung, bei der die Blattreihenfolge nicht gleichmäßig kreisförmig erfolgt, wird auch azyklisch genannt. Bei der wirteligen Anordnung spricht man auch von einer zyklischen Form, da die Blätter kreisförmig angeordnet sind.
Oftmals existiert auch eine Mischform aus der azyklischen und der zyklischen Form, bei der die äußeren Blätter azyklisch angeordnet sind, während die inneren Blätter eine wirtelige Stellung aufweisen. Dieser Übergang entsteht durch eine Unterbrechung der Blattbildung nach drei schraubig angeordneten Blättern.
Bei rein wirtelig angeordneten Blättern haben alle Blätter innerhalb des gleichen Kreisumlaufs den gleichen Abstand zueinander. Bei vielen Angiospermen-Arten existiert auch eine fixe Anzahl an Blättern pro Wirtel, wie bspw. bei der Tulpe. Außerdem stehen die Blätter des nächsten Wirtels dort, wo eine Lücke des vorherigen Wirtels herrscht. Dies lässt sich ebenfalls sehr gut bei der Tulpe beobachten.
Die Symmetrien von Blüten lassen sich in radiärsymmetrisch, disymmetrisch und monosymmetrisch zusammenfassen. Radiärsymmetrisch sind Blüten, sobald mehr als drei Symmetrieebenen durch die Blüte gelegt werden können, während disymmetrische Blüten nur zwei Symmetrieachsen besitzen. Nur eine Symmetrieachse weisen beispielsweise Lippenblütler wie Pfefferminze oder Lavendel auf.
Genetische Faktoren der Blütensymmetrie
Damit die Blüten ihre speziellen Symmetrien aufweisen können, werden verschiedene Gene exprimiert. Diese Gene werden auch als Blüten-Identitäts-Gene bezeichnet. Die wichtigsten dieser Gene sind FLC, SOC1, LFY und FT. FLC, auch Flowering Locus C genannt, ist nach der Vernalisation aktiv. Wird FLC deaktiviert, blüht die Pflanze zu früh.
SOC1 wird von FLC unterdrückt. Entfernt man die Wirkung des FLC, erfolgt dennoch kein Blühen, da hierfür die Aktivierung durch Gibberelline und Constans erfolgen muss. Constans ist ein Protein, das lichtgesteuert verschiedene Proteine exprimiert.
LFY wird ebenfalls durch eine lange Lichteinwirkung und Gibberelline aktiviert. Die Lichtaktivierung von LFY erfolgt jedoch über SOC1.
FT, der Flowering Locus T, wird durch eine lange Lichteinwirkung über Constans aktiviert. Gleichzeitig wird es durch FLC reprimiert (unterdrückt). Gleichzeitig aktiviert FT das Gen AP1.
LFY und AP1 sind Blütenmeristem-Identitäts-Gene. Zwischen ihnen besteht eine Wechselwirkung. AP1 wird zwar durch LFY aktiviert, dennoch können auch Blüten gebildet werden, wenn LFY deaktiviert wird, jedoch nicht mit den richtigen Organen.
Verschiedene Geschlechter von Blüten
Normalerweise weisen Angiospermen zwittrige (zweigeschlechtliche) Blüten auf. Allerdings existieren auch Arten, bei denen eingeschlechtliche Blüten ausgebildet werden. Hierbei wird zwischen männlichen und weiblichen Pflanzen unterschieden. Darüber hinaus können eingeschlechtliche Blüten entweder auf einer einzigen Pflanze sitzen oder getrennt auf verschiedenen.
Sitzen männliche und weibliche Blüten auf einer Pflanze, so wird dies als Monözie bezeichnet, während die Trennung von männlichen und weiblichen Blüten Diözie genannt wird. Weiterhin existieren auch polygame Pflanzen, die sowohl zwittrige als auch eingeschlechtliche Blüten aufweisen.
Die Auslösung der Blüte
Die Auslösung der Blüte ist ein Prozess innerhalb der Pflanze, der nicht umkehrbar ist. Er wird sowohl durch innere als auch durch äußere Faktoren beeinflusst. Derweil sind vier Signalwege bekannt, die eine Induktion der Blüte hervorrufen.
Genetische Faktoren
Im Laufe des Lebens einer Pflanze bildet sie an gewissen Zeitpunkten Blüten aus. Ohne die Auslösung durch äußere Faktoren wird dieser Prozess genetisch gesteuert, wie z.B. durch die Aktivierung des Gens FCA.
Vernalisation (Tageslänge)
Äußere Faktoren, die die Blüh-Induktion beeinflussen, sind die Tageslänge und die Temperatur. Um die Tageslänge feststellen zu können, besitzen Pflanzen verschiedene Photorezeptoren, wovon die Phytochrome und Cryptochrome die wichtigsten sind. Diese Systeme sind dazu in der Lage, ein Gen zu beeinflussen, das die Blüh-Induktion auslöst.
Vernalisation (Temperatur)
Pflanzen, die lediglich einmal blühen, benötigen eine intensive Kälteperiode, um Blüten zu bilden. Hierbei spielen einige Gene eine Rolle, die selbst die Induktion der Blüte vorantreiben, gleichzeitig aber das Gen FCA hemmen. Somit wird zwar die Blütenbildung ermöglicht, aber nicht ohne weitere Faktoren ausgelöst.
Hormonelle Faktoren
Eine bestimmte Klasse von Pflanzenhormonen, die Gibberelline, sind maßgeblich an der Blüh-Induktion beteiligt.
Bestäubung und Befruchtung von Blüten
Bestäubung
Wird bei Angiospermen der männliche Pollen auf die Narben übertragen, so spricht man von Bestäubung. Hierbei kann die Bestäubung innerhalb der gleichen Pflanze geschehen, aber auch zwischen zwei verschiedenen Pflanzen der gleichen Art. Die Selbstbestäubung wird auch als Autogamie bezeichnet, die Fremdbestäubung hingegen als Allogamie. Da die Autogamie zur Arterhaltung eher wenig förderlich ist, da hierbei kein genetisches Material ausgetauscht wird, versuchen Pflanzen diesen Prozess zu unterbinden.
Hierfür haben sich im Laufe der Evolution mehrere Prozesse herauskristallisiert, die Herkogamie und die Dichogamie.
Die Herkogamie beschreibt die räumliche Trennung von Androeceum und Gynoeceum, wodurch es für Bestäuber nahezu unmöglich ist, innerhalb einer Blüte die Bestäubung zu vollführen.
Andererseits ist es bei manchen Pflanzenarten auch üblich, dass eine zeitliche Trennung zwischen der Ausbildung von Androeceum und Gynoeceum existiert. Dieser Prozess wird Dichogamie genannt. Hierbei unterscheidet man zwischen vormännlichen und vorweiblichen Blüten. Allerdings verhindern Herkogamie und Dichogamie lediglich die Bestäubung innerhalb einer Blüte. Sie verhindern jedoch nicht die Bestäubung zwischen zwei Blüten der selben Pflanze. Deshalb gibt es noch die Selbstinkompatibilität, die durch genetische Faktoren eine Selbstbefruchtung verhindert.
Möglichkeiten der Bestäubung
Damit eine Bestäubung erfolgen kann, gibt es drei verschiedene Möglichkeiten, die sich Pflanzen zu Nutze machen. Sie können entweder über den Wind, das Wasser oder mithilfe von Tieren bestäubt werden. Je nach Art der Bestäubung haben sich weitere, markante Merkmale entwickelt.
Bei der Windbestäubung (Anemophilie) sind die Blüten meist nicht besonders ästhetisch, da sie keine Tiere anlocken müssen. Außerdem hängen die Blüten oft herab und bilden kein Nektar.
Für die Hydrophilie (Wasserbestäubung) bilden Pflanzen auch keine sonderlich ästhetischen Blüten aus, meist ist in den Früchten ein Luftgewebe enthalten, was das Schwimmen erleichtert.
Im Sinne der Zoophilie (Tierbestäubung) steht die Blume als bestäubungsbiologische Funktionseinheit im Vordergrund. Hierbei sind oftmals viele Blüten zu einer Blume zusammengefasst, wie es bspw. bei Gänseblümchen der Fall ist. Andererseits können in seltenen Fällen auch mehrere Blumen von einer Blüte gebildet werden.
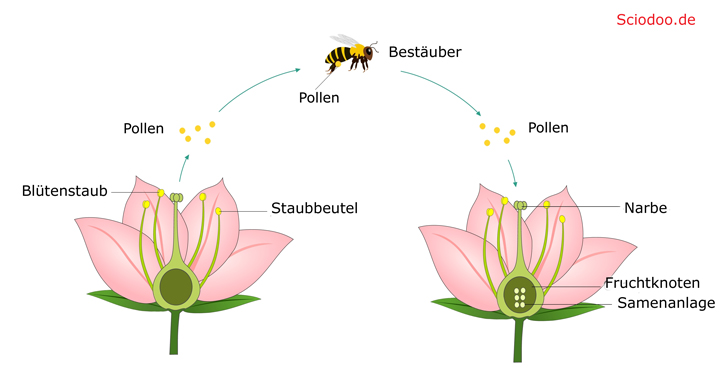
Bei jeder Bestäubung muss der männlichen Pollen von einer Blüte auf die andere Blüte gelangen, dort in die Narbe eindringen und den Fruchtknoten passieren. Dort findet die Verschmelzung zwischen Eizelle und Pollen statt, was als Befruchtung bezeichnet wird. Bei der Tierbestäubung werden Insekten durch Nektar, Blütenfarbe oder Geruch angelockt. Der Pollen bleibt an den Beinen der Bestäuber hängen und wird so auf die nächste Blüte übertragen.
Die meisten zoophilen Blüten sind zwittrig, haben eine ausgesprochene Farbvielfalt, riechen sehr stark und bilden Nektar, um Bestäuber anzulocken. Der Frauenschuh, eine Orchideenart, lockt Bestäuber in einen Kelch, aus dem der Bestäuber nicht herauskommt. Es sei denn, er klettert an der Rückseite des Kelches empor, wo er die Pollen auf die Narbe überträgt.
Befruchtung
Die Befruchtung erfolgt bei Angiospermen und Gymnospermen unterschiedlich. Bei den Gymnospermen (Nacktsamern) werden die Pollen zunächst auf die Mikropyle übertragen. Daraufhin werden sie in die Pollenkammer gezogen, nachdem der Bestäubungstropfen vertrocknet ist. Im weitern Verlauf werden entweder Spermatozoide freigesetzt oder das Auskeimen der Pollenschläuche erfolgt.
Bis die letztendliche Befruchtung erfolgt, können bei manchen Arten der Gymnospermen mehrere Monate vergehen.
Bei Angiospermen (Bedecktsamern) hingegen erfolgt zunächst die Bestäubung, indem die Pollen auf die Narbe des Stempels gebracht werden. Mithilfe eines Leitgewebes gelangt der Pollenschlauch von der Narbe zu den Samenanlagen.
Die letztendliche Befruchtung erfolgt, sobald die Pollenschläuche/Spermatozoide bei den Eizellen ankommen. Hierbei kommt es bei manchen Angiospermen zu einer doppelten Befruchtung, indem ein Spermakern mit der Eizelle verschmilzt, der andere hingegen mit dem Embryosackkern. Aus der verschmolzenen Eizelle entsteht daraufhin die Zygote, der verschmolzene Embryosackkern bildet das Nährgewebe für den Pflanzensamen.
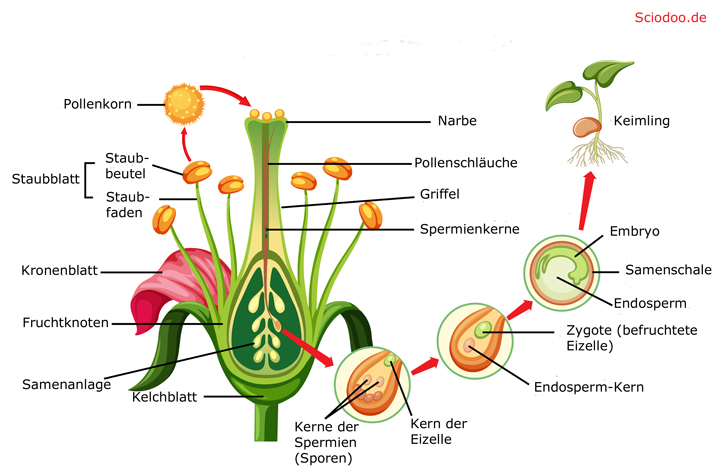
Fortpflanzung, Befruchtung und Reifung bei Pflanzen
Ist die Befruchtung erfolgt, so entwickelt sich die Zygote zum Embryo, während die Samenanlage zum Pflanzensamen und die Blüte zur Frucht heranwachsen.
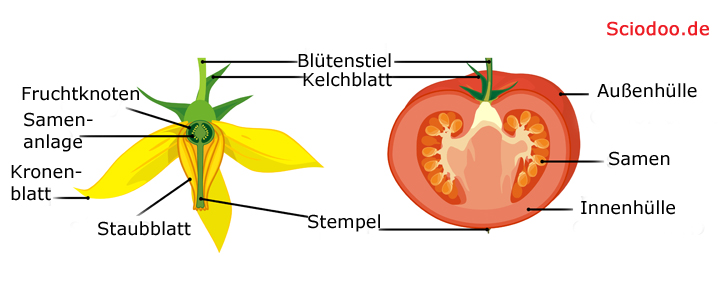
Die Blüte wird zur Frucht. Durch Geschmack werden Tiere angelockt, welche die Frucht fressen und den Samen irgendwo anders ausscheiden. Dort entsteht dann ein neue Keimling, welcher zur Jungpflanze heranwächst
Die Seneszenz der Blüten
Seneszenz beschreibt die Alterung von Zellen, Geweben oder Organen. Im Vergleich zum Tierreich werden bei Pflanzen die Sexualorgane (Blüten) ständig neu entwickelt. Dies liegt unter anderem daran, dass Blüten viel Energie und Nährstoffe verbrauchen, aber auch ein Ort zur Entstehung von Infektionen darstellen.
Außerdem würden bestäubte Blüten, die eventuell zur Befruchtung heranreifen, mit nicht bestäubten Blüten konkurrieren, was einen deutlichen Nachteil für die Pflanze mit sich bringen würde.
Damit insbesondere die Konkurrenz zwischen bereits bestäubten und nicht bestäuben Blüten ausbleibt, altern bestäubte Blüten sehr schnell. In vielen Pflanzen wird durch die Bestäubung das Hormon Ethylen freigesetzt, was wiederum dazu beiträgt, dass insbesondere die Kronenblätter verwelken. Wie beim Laubfall der Bäume geht auch die Rückbildung der Blüten mit einer Rückgewinnung von Nährstoffen einher.
Literatur
- Steffen Guido Fleischhauer (Autor), Roland Spiegelberger (Autor), Claudia Gassner (Autor), Blatt für Blatt: Über 800 Pflanzen nach Blattformen und Blüten einfach bestimmen. Pflanzen erkennen leicht gemacht – das praktische Bestimmungsbuch für das ganze Jahr, ISBN: 978-3039021758*
- Dieter Heß (Autor), Die Blüte: Struktur, Funktion, Ökologie, Evolution, ISBN: 978-3818600648*
Über den Autor
