Warum ist die Photosynthese eine endotherme Reaktion
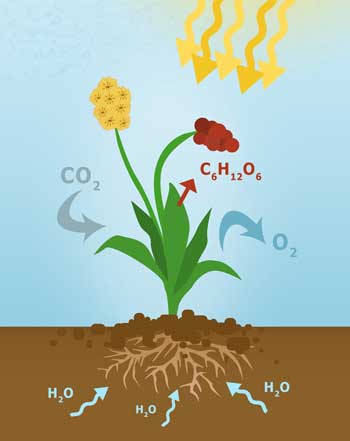
Die Photosynthese ist ein Stoffwechselprozess, welcher sich in verschiedene Teilprozesse unterteilt. Diese Teilprozesse sind bspw. die Lichtreaktion und die Dunkelreaktion, welche während der Fotosynthese isoliert ablaufen. Für diese Form des Stoffwechsels benötigt die Pflanze allerdings Energie von außen, welche als thermische Energie bzw. Wärmeenergie zur Verfügung steht. Im Winter, wenn diese Wärmeenergie fehlt, kann Fotosynthese – als eine endotherme Reaktion – deshalb weniger oder gar nicht stattfinden.
Inhalt
Was ist eine endotherme Reaktion
Chemische Reaktionen, denen zum Start Energie zugeführt werden muss und deren Standardenthalpiedifferenz positiv ist, werden als endotherm bezeichnet. Die Standardenthalpiedifferenz (ΔH) ist die Differenz zwischen zwei Wärmezuständen. Diese ergibt sich, indem man den Endzustand (HEndzustand) und den Ausgangszustand (HAusgangszustand) eines Systems, eines Objektes oder Teilchens – vor und nach der Reaktion – misst und dann die Differenz zwischen beiden errechnet.
Bei der Verbrennung ist der Wärmegehalt des Anfangszustandes (Ausgangsstoff + Feuer) höher als der Endzustand (verbrannter Stoff). Die Standardenthalpiedifferenz (ΔH = Endzustand – HAusgangszustand) ist negativ. Somit ist klar, dass bei dieser Reaktion eine entsprechende Wärme abgegeben wurde. Man bezeichnet Reaktionen, bei denen mehr Energie freigesetzt als zugeführt wurde, als exotherme Reaktion.
Das Gegenstück zu den exothermen sind die endothermen Reaktionen, bei denen Wärme während der Reaktion aufgenommen wird. Dadurch ist der Wärmegehalt des Endzustandes (HEndzustand) größer als beim Anfangszustand (HAusgangszustand), wodurch der Standardenthalpiedifferenz (ΔH) positiv wird. Halten wir fest, dass bei endothermen Reaktionen weniger Energie freigesetzt wird, als aufgenommen wurde. Die Energie verbleibt somit im System.
Was bedeutet Enthalpie (H) bei endothermen Reaktionen?
Der Wärmegehalt bzw. wissenschaftlich als Enthalpie (H) bezeichnet, wird zur Berechnung von endothermen Reaktionen herangezogen. Diese Größe ist nicht direkt messbar, sondern wird mathematisch als Summe der inneren Energie (U) eines Systems mit dem Produkt von Druck (p) und Volumen (V) ausgedrückt (H = U + p*V).
Da Wärme grundsätzlich nichts anderes ist als die kinetische Energie (Bewegungsenergie) eines Objekts ist, kann die Enthalpie als Wärmegehalt eines Objekts bei gleichbleibendem Druck angesehen werden. Der Druck (p) muss bei dieser Betrachtung konstant bleiben, da sich bei veränderten Druck auch die Bewegungsintensität der Teilchen eines Objektes ändern. Durch Wärme wird die Bewegungsintensität ebenfalls angeregt, weshalb der Druck fixiert und als Konstante einfließen muss – um klare Ergebnisse erzielen zu können.
Um zu berechnen, ob eine Reaktion endotherm abläuft, wird die Differenz zwischen den Enthalpien, also den Produkten und den Edukten (Ausgangsstoffen), gebildet (HEnd – HAus).
Ist diese Differenz größer als 0, so handelt es sich um eine endotherme Reaktion. Somit sind sämtliche Reaktionen, denen zu Beginn mehr Energie zugeführt werden muss, als schlussendlich frei wird, endotherme Reaktionen. Wird jedoch mehr Energie bspw. in Form von Wärme freigesetzt, als anfangs zugeführt wurde, bezeichnet man diese Reaktionen als exotherm.
Als Beispiel einer rein physikalischen Reaktion, die endotherm abläuft, lässt sich das Schmelzen von Eis heranziehen. Da hierbei keine Stoffumwandlung im klassischen Sinne, sondern nur eine Aggregatszustandsänderung stattfindet, ist dies keine chemische Reaktion. Diese Reaktion verläuft endotherm, da der Ausgangsstoff (Eis) einen niedrigeren Wärmegehalt als der Endstoff (Wasser) hat, weshalb äußerliche Wärme den Schmelzvorgang begleitet bzw. anstößt. Der Standardenthalpiedifferenz (ΔH) ist hierbei positiv.
Ein klassisches Beispiel für eine exotherme Reaktion ist die Thermit-Reaktion. Diese Reaktion von Eisen(III)-Oxid und Aluminium zu Eisen und Aluminiumoxid setzt Temperaturen von ca. 2000°C frei. Die Photosynthese ist hingegen eine endotherme Reaktion, welche externe Energie (Sonnenlicht) benötigt und die über mehrere Teilreaktionen abläuft, welche immer wieder die Lichtstrahlen der Sonne als Energiequelle nutzen.
Die Teilreaktionen der Photosynthese
Die Photosynthese ist der Stoffwechselweg, den Algen, einige Bakterien, Moose, Farne und Pflanzen verwenden, um lebensnotwendige Kohlenhydrate wie z.B. Glukose aufzubauen. Sie lässt sich in zwei Teilreaktionen gliedern, denen wiederum weitere Reaktionen innewohnen. Die beiden Teilreaktionen werden als Licht- und Dunkelreaktion bezeichnet. Die Dunkelreaktion kann jedoch nicht ohne die vorangegangene Lichtreaktion stattfinden, da hierbei Produkte generiert werden, die während der Dunkelreaktion weiterverarbeitet werden. Weiterhin lässt sich die Photosynthese in die oxygene und die anoxygene Photosynthese unterteilen, je nachdem ob als Endprodukt molekularer Sauerstoff oder andere anorganische Moleküle gebildet werden.
Lichtreaktion während der Photosynthese
Die lichtabhängige Reaktion (Lichtreaktion) finden in den Chloroplasten der Pflanzenzelle statt. Innerhalb der Chloroplasten existieren kleine Untereinheiten, die sog. Thylakoide. In den Thylakoidmembranen sind die für die Lichtreaktion notwendigen Pigmente eingelagert. Zu diesen Pigmenten zählt u.a. auch das Chlorophyll. Aber auch Carotinoide oder Phycobiline zählen zu diesen Pigmenten.
Während die Lichtreaktion an der Thylakoidmembran stattfindet, geschieht die nachgeschaltete lichtunabhängige Reaktion innerhalb des Stromas (der flüssige Raum innerhalb des Chloroplasten). Der grundlegende Prozess der Lichtreaktion ist die Umwandlung von Lichtenergie in chemische Energie. Die chemische Energie wird hierbei in den Molekülen ATP und NADPH gespeichert und kann somit der Dunkelreaktion zur Verfügung gestellt werden.
Der molekulare Ablauf der Lichtreaktion
Die Energie des Lichts ist in den sog. Photonen gespeichert und kann in Form von Wärme oder chemischer Energie freigegeben werden. Die oben beschriebenen Pigmentmoleküle können die Lichtenergie der Photonen nutzen, um diese in chemische Energie umzuwandeln. Als Beispiel soll hierbei das Chlorophyll dienen. Dieses absorbiert die Energie des Photons und wird dadurch von seinem energetischen Grundzustand in einen Zustand höherer Energie versetzt. Es wird „angeregt“.
Diese Umwandlung der Energie liegt im ersten Hauptsatz der Thermodynamik begründet, da dieser besagt, dass Energie weder vernichtet noch erschaffen werden kann. Somit ist die im Photon enthaltene Energie lediglich in der angeregten Form des Chlorophylls gespeichert und ist nicht verlorengegangen. Im Idealfall entspricht somit die Energiezunahme des angeregten Chlorophylls exakt der Energie, die das Photon in sich trug, sofern keine Verluste in Form von Wärme o.ä. stattgefunden haben. Auf atomarer Ebene bedeutet die Anregung des Chlorophylls einen Elektronentransport von einer inneren Elektronenschale auf eine äußere Elektronenschale. Auf das „gesprungene“ Elektron wirken nun weniger Anziehungskräfte des Atomkerns, weshalb es reaktionsfreudiger wird.
Räumlich findet diese Reaktion in den Pigmenten statt, die in den Thylakoidmembranen angeordnet sind. Zur Optimierung der Lichtaufnahme liegen diese Pigmente in sog. Antennenkomplexen vor. Die Gesamtheit der Antennenkomplexe liegt in Intermembranproteinen, den sog. Fotosystemen vor. Innerhalb der Fotosysteme existieren Reaktionszentren, die die gesamte Lichtenergie bündeln.
Ein Reaktionszentrum besteht wiederum aus zwei Chlorophyll-a-Molekülen. Das angeregte Elektron eines Chlorophyllmoleküls des Antennenkomplexes wandert zu dem Reaktionszentrum und wird von dort aus auf ein Akzeptormolekül übertragen. Das Akzeptormolekül reicht das Elektron innerhalb der Thylakoidmembran weiter. Da das Chlorophyll, das das Elektron auf das Akzeptormolekül überträgt, eine negative Ladung in Form eines Elektrons verliert, spricht man hierbei von einer Oxidation. Das Fotosystem II, an dem diese Reaktion stattfindet, wird auch als P680 bezeichnet, da die Wellenlänge des Lichts, die optimal absorbiert werden kann, 680 Nanometer beträgt.
Während der Oxidation des Chlorophylls entsteht eine sog. Elektronenlücke, wodurch das Chlorophyll in einen instabilen Zustand gerät und versucht, ein Elektron aufzunehmen. Die zum Füllen der Elektronenlücke benötigten Elektronen stammen aus dem Wasser, das für die Photosynthese ebenfalls benötigt wird. Das Wasser wird an der Thylakoidmembran an einem wasserspaltenden Komplex in die Bestandteile Sauerstoff und Wasserstoff gespalten. Der Wasserstoff liegt jedoch in Form von Protonen und nicht als Molekül vor. Der Sauerstoff eines Wassermoleküls reagiert mit einem anderen Sauerstoffatom zu molekularem Sauerstoff (O2) und wird abtransportiert.
Neben den Protonen liegen jedoch weitere Elektronen vor, die während der Wasserspaltung entstanden sind. Eines dieser Elektronen kann nun die Elektronenlücke im Fotosystem II auffüllen. Daraus lässt sich auch ableiten, dass der während der Photosynthese gebildete Sauerstoff aus dem Wasser stammt und nicht etwa aus dem aus der Luft aufgenommenen Kohlendioxid (CO2).
Das Elektron, das vom angeregten Chlorophyll des Reaktionszentrums auf das Akzeptormolekül übertragen wurde, wandert während der Membranpassage über verschiedene Proteinkomplexe. Dazu zählen das Plastochinon, das Cytochrom und das Plastocyanin. Diese Proteinkomplexe bilden die Grundlage für die Elektronentransportkette, um die Photosynthese am folgenden Fotosystem I (P700) weiterhin betreiben zu können.
Insbesondere am Cytochrom wird die Energie der Elektronen dazu genutzt, um Protonen aus dem Stroma in das Thylakoidlumen (Thylakoid-Innenraum) zu transportieren. Zusammen mit den Protonen, die während der Wasserspaltung entstanden sind, wird somit ein Konzentrationsgradient der Protonen im Inneren der Thylakoide aufgebaut. Aufgrund der positiven elektrischen Ladung der Protonen entsteht neben dem Konzentrationsgradienten auch ein elektrischer Gradient, der dazu führt, dass das Stroma negativer geladen ist als das Thylakoidlumen.
Der zusammenfassend als elektrochemischer Gradient bezeichnete Gradient beinhaltet die Protonenmotorische Kraft, die eine weitere Form der Energie beinhaltet, um weitere Reaktionen ablaufen lassen zu können. Da die Thylakoidmembran für Protonen grundsätzlich undurchlässig ist, wird ein Kanal benötigt, um die Protonen vom Thylakoidlumen in das Stroma zu transportieren. Dieser Kanal wird als ATP-Synthase bezeichnet.
Die ATP-Synthase koppelt den Protonentransport an die Synthese von ATP (Adenosintriphosphat). Hierbei wird mithilfe der protonenmotorischen Kraft auf enzymatische Weise das Adenosindiphosphat (ADP) mit einem Phosphatrest zu ATP verknüpft. Dieser Prozess wird auch als Fotophosphorylierung bezeichnet. Die Fotophosphorylierung ist eine endotherme Reaktion. Ohne die Fotophosphorylierung würde der Protonengradient stetig ansteigen und nicht abgebaut werden können.
Das Elektron, das ursprünglich vom Fotosystem II über die Proteinkomplexe Plastochinon, Cytochrom und Plastocyanin gewandert ist, erreicht das Fotosystem I (P700). Auch hier wird Licht absorbiert, was wiederum zu einer Elektronenübertragung im Reaktionszentrum auf ein Akzeptormolekül führt. Auch hierbei entsteht eine Elektronenlücke, die wiederum von den Elektronen aus dem Elektronentransport vom Fotosystem II aufgefüllt wird.
Nachdem das Elektron im Fotosystem I vom Akzeptormolekül aufgenommen wurde, wird es zunächst auf einen Proteinkomplex übertragen, der Ferredoxin genannt wird. Daran angeschlossen ist die NADP+-Reduktase. Die NADP+-Reduktase ist ein Enzym, das insgesamt zwei Elektronen, zwei Protonen und ein Molekül NADP+ benötigt, um daraus den Wasserstoffüberträger NADPH+H+ zu generieren.
Zwischenfazit der Lichtreaktion
Der Elektronentransport lässt sich folgendermaßen zusammenfassen: Zunächst werden Chlorophyllmoleküle im Fotosystem II angeregt, die ein Elektron an das Reaktionszentrum weiterleiten. Von dort aus wird das Elektron auf ein Akzeptormolekül übertragen und wandert im Anschluss über Plastochinon, Cytochrom und Plastocyanin zum Fotosystem I.
Die entstandene Elektronenlücke im Fotosystem II wird durch Elektronen gefüllt, die während der photolytischen Spaltung des Wassers entstanden. Im Fotosystem I erfolgt wieder eine Anregung des Chlorophylls inkl. einer Elektronenweitergabe an ein Akzeptormolekül. Von dort aus wandert das Elektron über Ferredoxin an die NADP+-Reduktase, wobei aus zwei Elektronen, zwei Protonen und einem NADP+ das Reduktionsäquivalent NADPH+H+ gebildet wird. Die hierfür benötigten Protonen stammen zum einen aus der Wasserspaltung, zum anderen aus dem Elektronentransport über das Cytochrom.
In beiden Fällen wird ein Protonengradient im Inneren der Thylakoide aufgebaut, der über die ATP-Synthase abgebaut wird, wodurch die Protonen letztendlich der NADP+-Reduktase zur Verfügung stehen. Da während des Elektronentransports der Lichtreaktion ATP und NADPH+H+ gebildet wurde, kann die Dunkelreaktion ablaufen.
Dunkelreaktion während der Photosynthese
Die Dunkelreaktion bzw. lichtunabhängige Reaktion der Photosynthese wird auch als Calvin-Zyklus bezeichnet. Dieser Prozess findet, wie die lichtabhängige Reaktion, in allen photosynthetisch aktiven Organismen statt. Das Ziel des Calvin-Zyklus ist die Synthese von Glukose, welche für verschiedene Prozesse wie z.B. das Wachstum benötigt wird. Andererseits kann in weiteren Schritten während der Zellatmung die Glukose auch abgebaut werden, um erneut Energie freizusetzen.
Wie eingangs erwähnt, kann der Calvin-Zyklus nur dann ablaufen, sofern während der Lichtreaktion genug ATP und NADPH gebildet wurde. Die Bezeichnung als Dunkelreaktion bzw. lichtunabhängige Reaktion kann deshalb missverstanden werden. Grundsätzlich kann diese Reaktion zwar ohne die direkte Einwirkung des Lichts ablaufen, doch damit diese Reaktion ablaufen kann, werden die Produkte der lichtabhängigen Reaktion benötigt, wodurch diese Reaktion indirekt lichtabhängig ist. Darüber hinaus existieren jedoch Ausnahmen der Photosynthese, bei denen der Calvin-Zyklus komplett autonom abläuft.
Der molekulare Ablauf während der Dunkelreaktion
Der Ort des Calvin-Zyklus findet im Stroma der Chloroplasten statt. Grob lässt sich der Calvin-Zyklus in drei Phasen untergliedern. Diese sind:
- CO2-Fixierung,
- die Reduktionsphase
- und die Regenerationsphase des CO2-Akzeptors.
Während der CO2-Fixierung wird das CO2, das über die Stomata (Spaltöffnungen) der Blätter einströmt, fixiert, damit kein Ausstrom erfolgt und die Synthese der Glukose beginnen kann. Die Fixierung des CO2 wird mithilfe des Moleküls Ribulose-1,5-bisphosphat (RuBP) absolviert.
Im ersten Reaktionsschritt wird das Kohlenstoffdioxid auf RuBP übertragen. Diese Reaktion wird durch das Enzym Ribulosebisphosphat-Carboxylase-Oxygenase (Rubisco) katalysiert. Rubisco ist das auf der Erde am häufigsten vorkommende Enzym. Bei der Fixierung des Kohlenstoffdioxids (C1-Körper, da nur ein Kohlenstoffatom enthalten ist) an dem C5-Körper RuBP entsteht zunächst ein extrem instabiler C6-Körper, der nahezu sofort in zwei C3-Körper zerfällt. Die beiden C3-Körper heißen 3-Phosphoglycerat bzw. 3-Phosphoglycerinsäure (3PG).
Während der zweiten Phase, der Reduktionsphase, wird aus diesen beiden Molekülen 3PG ein Kohlenhydrat, das sog. Glycerinaldehyd-3-Phosphat (G3P) synthetisiert. Dieser Prozess ist endergonisch bzw. endotherm, da ihm Energie zugeführt werden muss. Diese Energie wird während der ersten Teilreaktion der Reduktionsphase in Form von ATP bereitgestellt, das während der Lichtreaktion gebildet wurde.
Damit Energie aus dem ATP frei wird, muss ein Phosphatrest abgespalten werden, wodurch ADP (Adenosindiphosphat) entsteht. Dieser Phosphatrest kann nun auf 3-Phosphoglycerat übertragen werden. Die Übertragung eines Phosphatrests wird auch Phosphorylierung genannt. Da das 3-Phosphoglycerat nach der Aufnahme des Phosphatrests über zwei Phosphatgruppen verfügt, heißt diese Verbindung 1,3-Bisphosphoglycerat (1,3-BPG).
Im zweiten Teilschritt der Reduktionsphase, in dem aus 1,3-Bisphosphoglycerat das Glycerinaldehyd-3-Phosphat gebildet wird, wird das ebenfalls in der Lichtreaktion gebildete NADPH+H+ als Elektronenüberträger benötigt. Das NADPH+H+ überträgt zwei Elektronen und ein Proton. Gleichzeitig wird eine Phosphatgruppe des 1,3-Bisphosphoglycerats mit einem Sauerstoffatom abgespalten. Das Sauerstoffatom geht eine Verbindung mit zwei Protonen ein, wodurch Wasser entsteht, und wird aus dem Calvin-Zyklus ausgeschleust.
Durch die Reduktion des 1,3-Bisphosphoglycerats entsteht eine chemische Veränderung. Aus der endständigen Carboxylatgruppe des 1,3-BPG wird eine endständige Aldehydgruppe am G3P. Da es sich bei dieser Reaktion um eine Redoxreaktion handelt, geht mit der Reduktion des 1,3-BPG die Oxidation des NADPH+H+ einher. Die Moleküle ADP und NADP+ stehen nun wieder für die Lichtreaktion zur Verfügung.
Im folgenden Reaktionsschritt können zwei Möglichkeiten auftreten. Entweder wird das G3P zur weiteren Synthese von Zuckern eingesetzt, oder die Regeneration des CO2-Akzeptors RuBP wird vorangetrieben. Die Synthese von Zuckern läuft in etwa 1/6 der Fälle ab, während die Regeneration zu 5/6 stattfindet. Die Verhältnisse beziehen sich hierbei auf das Mengenverhältnis an G3P, das genutzt wird. Die Synthese von Zuckern geschieht durch die Abspaltung des Phosphatrests zweier Moleküle G3P. Hierdurch entstehen zwei sog. Triosen (Zucker mit 3 C-Atomen).
Werden diese beiden Triosen miteinander verknüpft, entsteht eine Hexose (Zucker mit 6 C-Atomen) wie z.B Glukose oder Fructose. Diese Zucker können zu deutlich größeren Netzwerken verknüpft werden, wie z.B. Stärke, Cellulose oder Hemicellulose. Die Regenerationsphase gliedert sich in viele komplexe Reaktionen, die zunächst die Bildung des Moleküls Ribulose-Monophosphat (RuMP) zum Ziel haben. Durch eine erneute Spaltung des Moleküls ATP wird ein Phosphatrest frei, der auf RuMP übertragen wird, wodurch Ribulose-1,5-Bisphosphat entsteht.
Zwischenfazit der Dunkelreaktion
Da die Glukose bzw. Fruktose aus 6 Kohlenstoffatomen besteht und pro Durchlauf des Calvin-Zyklus ein CO2-Molekül verwendet wird, muss der Calvin-Zyklus insgesamt 6 Mal durchlaufen werden, um ein Molekül Glukose zu generieren. Hierfür werden insgesamt 18 Moleküle ATP und 12 Moleküle NADPH+H+ benötigt.
Die Summengleichung sieht demnach folgendermaßen aus: 6 CO2 + 12 H2O + 12 NADPH+H+ + 18 ATP -> C6H12O6 + 6 O2 + 6 H2O + 18 ADP + 18 Pi
Der Begriff der lichtunabhängigen Reaktion ist etwas irreführend, da hierfür Produkte der lichtabhängigen Reaktion benötigt werden. Darüber hinaus wird der Calvin-Zyklus durch Licht angeregt, indem der pH-Wert des Stromas durch Licht verändert wird. Diese pH-Wert-Änderung ermöglicht erst die Aktivität vieler Enzyme, die am Calvin-Zyklus beteiligt sind. Darüber hinaus ist auch der lichtinduzierte Elektronentransport ein Indiz dafür, dass der Calvin-Zyklus nicht komplett lichtunabhängig ist. Dieser Elektronentransport bewirkt eine strukturelle Veränderung der Enzyme, die daraufhin aktiviert werden.
Zusammenfassung
- Die Photosynthese ist eine endotherme Reaktion. Der Vorgang benötigt demnach eine Energiequelle, welche die Startenergie bereitstellt.
- Auch sämtliche Teilreaktionen der Photosynthese sind endotherme Reaktionen, wenngleich die Hydrolyse von ATP eine exotherme Reaktion darstellt. Doch die gesamte Bilanz der Photosynthese weist darauf hin, dass es sich um eine endotherme Reaktion handelt.
- Als Beispiel sei hier die Fotophosphorylierung genannt, durch die der Protonengradient indirekt abgebaut wird.
- Aber auch die Synthese von ATP aus ADP und das anorganische Phosphat Pi ist eine endotherme Reaktion, genauso wie die Synthese von NADPH+H+.
- Auch die Synthese des Zuckers während des Calvin-Zyklus ist eine endotherme Reaktion. Grundsätzlich sind mehr Teilreaktionen der Photosynthese endotherm als exotherm, was zu einer insgesamt endothermen Reaktion führt.
- Weiterhin ist die lichtunabhängige Reaktion der Photosynthese nicht so lichtunabhängig wie man vermuten mag. Einerseits ist der Calvin-Zyklus auf die Produkte der lichtabhängigen Reaktion angewiesen, andererseits finden weitere lichtinduzierte Prozesse statt, die den Calvin-Zyklus auf enzymatischer Eben erst ermöglichen.
- Somit bleibt die wichtigste Energiequelle der Photosynthese das Licht, um alle endothermen Teilreaktionen sowohl direkt als auch indirekt voranzutreiben. Aus diesem Grund findet im Winter weniger oder gar keine Photosynthese statt.
Über den Autor
