Grundorgane einer Blütenpflanze, Samenpflanze: Aufbau und Struktur
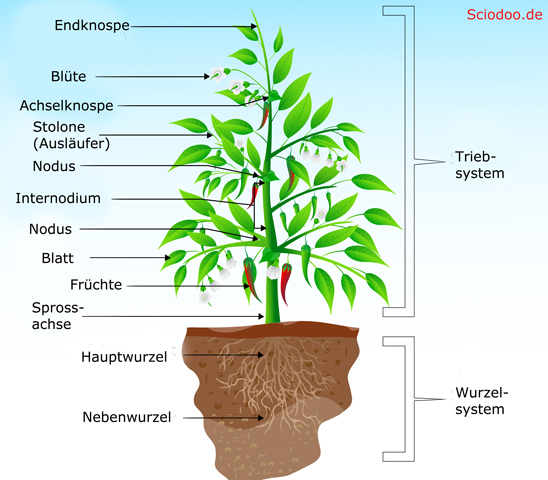
Die Grundorgane eine Samenpflanze sind Blatt, Wurzel und Sprossachse. Die Blüte ist ein weiteres Organ von Blütenpflanzen.
Samen- bzw. Blütenpflanzen existieren bereits seit mehreren Millionen Jahren und stellen immer noch eine große Gruppe im Reich der Pflanzen dar. Ihre Vielfalt ist äußerst komplex, ebenso die zugrundeliegenden anatomischen Merkmale. Um Unterschiede und Gemeinsamkeiten zwischen verschiedenen Pflanzen besser verstehen zu können, ist es hilfreich, sich mit den einzelnen Merkmalen und dem allgemeinen Aufbau von Samenpflanzen auseinanderzusetzen.
Inhalt
Was sind Blütenpflanzen
Samenpflanzen, die wissenschaftlich als Spermatophytina bezeichnet werden, stellen eine Gruppe im Reich der Pflanzen dar. Da sie zur Fortpflanzung zunächst Blüten bilden, aus denen dann Samen entstehen, werden sie oftmals als Blütenpflanzen bezeichnet. Weiterhin kann man die Samenpflanzen in Bedecktsamer (Angiospermen) und Nacktsamer (Gymnospermen) untergliedern. Die Bedecktsamer sind auch unter der Bezeichnung Magnoliopsida bekannt.
Weiterhin lassen sich Samenpflanzen auch in Monokotyle und Dikotyle Pflanzen unterscheiden. Die Bezeichnung steht für das Vorhandensein eines oder zweier Keimblätter (mono = eins, di = zwei) nach Auskeimen eines Samens.
Welche Pflanzen gehören zu den Blütenpflanzen
Man kann annehmen, dass die Begrifflichkeit lediglich Blumen oder andere Pflanzen – welche für ihre Blüten bekannt sind, einbezieht. Das reicht allerdings nicht aus. Zu den Blüten- bzw. Samenpflanzen gehören fast alle höheren Pflanzen, welche man wissenschaftlich auch als Gefäßpflanzen bezeichnet.
Abzugrenzen sind von den Gefäßpflanzen die Moose, Flechten und andere niedere Pflanzen (Kryptogame). Historisch zählte man in der Botanik auch Algen, Pilze und Bakterien zu den niederen Pflanzen. Heute weiß man, dass Algen, Pilze und Bakterien keine Pflanzenzellen besitzen, weshalb diese in jeweils eigene Reiche und Domänen eingeordnet werden.
Gefäßpflanzen, deren sexuelle Fortpflanzung ohne Blüte auskommen, sind Farne und Bärlapppflanzen. Alle andere Pflanzen, also alle Bäume, sämtliche Sträucher, Gräser und Kräuter – sind Samenpflanzen. Allerdings werden Nacktsamer, wie bspw. Nadelbäume, welche keine echten Blüten austreiben – nicht als Blütenpflanzen betrachtet. In der Wissenschaft sind demnach alle Bedecktsamer gleichzusetzen mit den Blütenpflanzen, deren Grundorgane aber – bis auf die Blüte – mit den Nacktsamern übereinstimmen.
Ein Unterscheidungsmerkmal von Samenpflanzen zu niederen Pflanzen ist unter anderem die klare Trennung in bestimmte Pflanzenorgane, die es so bei niederen Pflanzen nicht gibt. Und diese sogenannten Grundorgane einer Blütenpflanze schauen wir uns jetzt näher an.
Die Bedeutung der Grundorgane von Samenpflanzen
Bevor wir uns den Aufbau und die Anatomie der vier Grundorgane (Blatt, Sprossachse, Wurzel, Blüte) genauer anschauen, will ich kurz deren Bedeutung für die Pflanze erläutern.
Die Blüte ist das Anlockorgan aller Blütenpflanzen, welches die Fortpflanzungsorgane (Fruchtblatt und Staubblatt) ausbildet. Die Staubblätter bilden die Pollen, welche im Zuge einer Bestäubung durch Wind oder Insekten auf die Fruchtblätter gelangen.

Insekten dienen als Bestäuber, indem sie Pollen verbreiten
Im Fruchtblatt befindet sich die Samenanlage mit der Eizelle, welche befruchtet wird. Durch die Befruchtung verschmelzen Ei– und Samenzelle und es entsteht der Pflanzenembryo. Die Samenanlage, in welcher die Eizelle eingelagert war, entwickelt sich nun weiter zum Pflanzensamen und die Blüte wird zur Frucht. Diese Frucht (z.B. Obst) reift heran, fällt ab und wird von Tieren gefressen. Der Kot der Tiere wird irgendwo ausgeschieden und enthält dann den Pflanzensamen, welcher im richtigen Milieu zum Keimling heranreift.
Der Keimling benötigt Energie, um eine Jungpflanze zu werden. Diese Energie gewinnt der Keimling und auch die spätere Pflanze aus ihrem Stoffwechsel. Der Stoffwechselweg der Pflanzen ist die Photosynthese, bei der aus Wasser und Kohlendioxid ein Pflanzenzucker hergestellt wird. Dieser Pflanzenzucker ist der körpereigene Betriebsstoff des pflanzlichen Organismus. Es handelt sich um ein Kohlenhydrat, also einen Energieträger, dessen Energie die Pflanze für Wachstum, Entwicklung oder Fortpflanzung investieren kann.
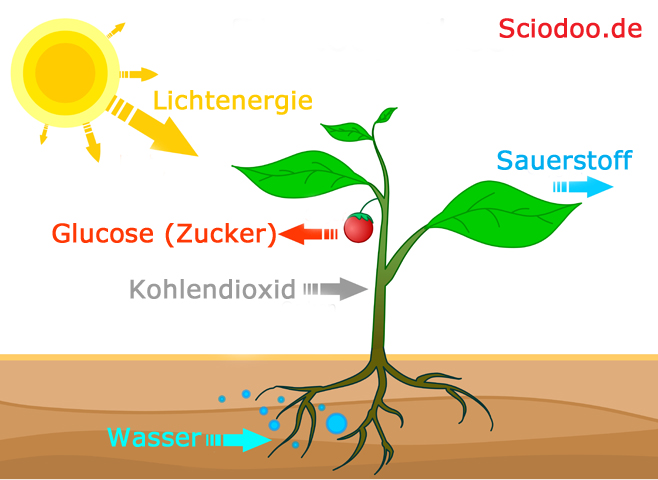
Bei der Photosynthese wird durch den Einsatz von Lichtenergie aus Kohlendioxid + Wasser ein Zucker und Sauerstoff hergestellt
Wie geht’s weiter?
Jeder Pflanze ist ein Organismus. Und wie jedes Lebewesen besteht auch die Pflanze aus Zellen. Die gewonnenen Energie aus den hergestellten Kohlenhydraten investiert die Pflanze ins Zellwachstum, wo sogenannte Zellorganelle aufgebaut werden. Jede Pflanzenzelle besteht aus Zellorganellen, wie bspw. Chloroplasten, Cytoplasma, Zellwand, Mitochondrium oder Zellkern. Dabei erfüllt jedes Organell eine bestimmte Aufgabe für die Zelle und somit für die Pflanze. Ein Hauptbestandteil dieser Zellorganellen sind Proteine.
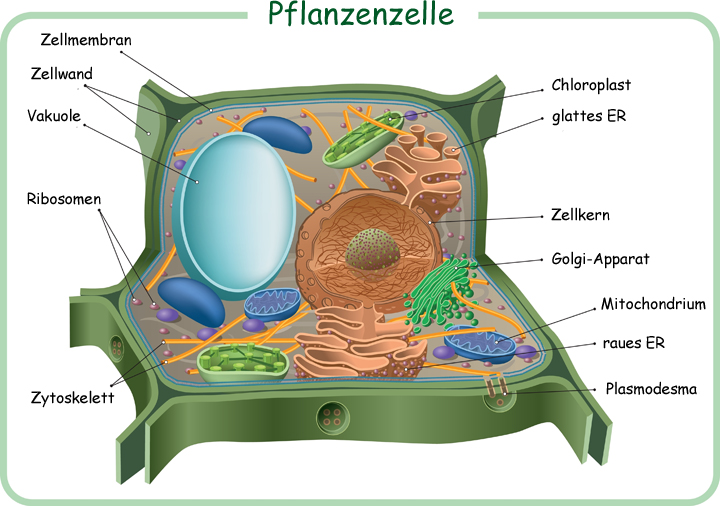
Jeder Teil einer Pflanze besteht aus Zellen, welche wiederum aus Organellen bestehen
Und diese Proteine muss die Pflanze ebenfalls aufbauen. Somit baut sie Kohlenhydrate für den Energiestoffwechsel auf und Proteine, um Gewebestrukturen zu schaffen. Und für den Aufbau dieser Proteine benötigt die Pflanze verschiedene Atome, welche sie über die Wurzel aufnimmt. Denn im Erdreich existieren sogenannte Nährsalze, wie Nitrate, Phosphate oder Sulfate.
Indem die Pflanze diese Atome aufnimmt und neu ordnet, baut sie ihre eigenen Proteine und somit Gewebestrukturen auf. Dazu investiert die Pflanze die Energie – indem sie den Zucker wieder abbaut. Außerdem entsteht bei der Photosynthese auch Sauerstoff. Dieser wird ebenfalls benötigt. Denn Sauerstoff wird als Reaktionspartner für diverse chemische Reaktionen gebraucht, die beim Proteinaufbau bzw. der Atom-Neuordnung nötig sind.
Die Pflanze nimmt demnach Nährstoffe über die Wurzel auf. Da die Partikel viel zu groß sind, um in die Zellen der Pflanze zu gelangen, müssen diese gelöst werden. Das bedeutet, dass diese in Wasser aufgelöst auftreten müssen, um durch die Zellmembran der Pflanzenzelle zu gelangen. Im Wasser werden die gelösten Nährstoffe durch die Pflanze transportiert und erreichen so jede Zielzelle.
Wie kommt das Wasser bis ganz nach oben?
Um genügend Druck aufbauen zu können, besitzen die Blätter kleine Öffnungen (Stoma). Diese Spaltöffnungen lassen Wasser verdunsten, wodurch ein Innendruck in der Pflanze entsteht – welcher das Wasser bis nach oben pumpen lässt.
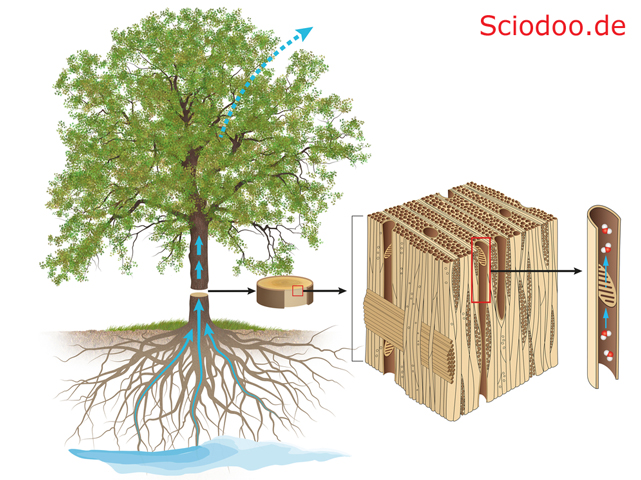
Durch Verdunstung an den Blättern entsteht ein Druck, wodurch Wasser von der Wurzel bis zur Baumkrone transportiert werden kann
Aber die Blätter der Pflanze sind nicht nur für den Druck zuständig. Im Blattgrün befinden sich die Chloroplasten, welche Sonnenlicht in einer ganz bestimmten Wellenlänge absorbieren können. Durch die Absorbierung nimmt die Pflanze Wärmeenergie von außen auf, welche sie dann in die Photosynthese (Stoffwechsel) als Startenergie investiert.
Halten wir fest….
Durch die Photosynthese wird Zucker (wie Glucose) aufgebaut, welcher Energie liefert. Diese Energie investiert die Pflanzen in Lebensprozesse. Für den Aufbau der Zellen benötigt die Pflanze bestimmte Nährsalze, welche im Erdboden vorkommen. Durch Wasser werden diese Nährstoffe gelöst, können in der Pflanze transportiert werden. Die Wurzeln nehmen Wasser und Nährstoffe auf. Durch die Spaltöffnungen an den Blättern wird Wasser ausgeschieden, wodurch sich der Wasserdruck in der Pflanze erhöht und weiteres Nährstoffwasser bis nach oben pumpen lässt. Die Sprossachse ist das Verbindungsstück zwischen Blatt und Wurzel. Und die Blüten sind Organe, welche der Fortpflanzung dienen.
Aufbau einer Samen- bzw. Blütenpflanze
Der grundlegende Aufbau einer Samenpflanze lässt sich in vier Bereiche gliedern. Diese sind der Wurzelbereich, die Sprossachse (bei Bäumen der Stamm), die Blätter und die Blüten.
Plasmodesmos als Verbindung zwischen zwei Nachbarzellen
Ein Plasmodesmos bzw. Plasmodesma ist ein dünner schlauchförmiger Plasmastrang zwischen zwei benachbarten Pflanzenzellen. Die Verbindung geht durch die Zellwand zweier Zellen hindurch und schafft so einen Stoffaustausch über die Zellgrenze hinweg.
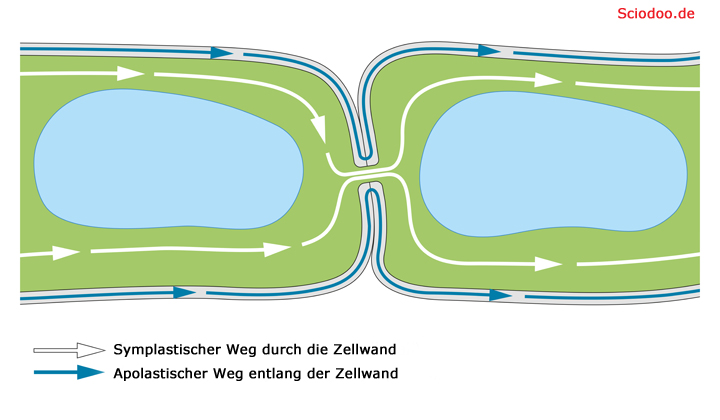
Symplast und Apoplast in einer Pflanzenzelle
Die Gesamtheit aller Plasmodesmen-Verbindungen einer Pflanze wird als Symplast bezeichnet. Dieses betrifft das Zellinnere. Die Gesamtheit aller Zellwände und aller Zwischenzellräume wird als Apoplast bezeichnet.
Der Wurzelbereich

Xylem ist das Leitbündelsystem, welches Wasser und darin gelöste Nährstoffe von der Wurzel in die Pflanze verteilt. Das Phloem, als Leitungssystem, verteilt Stoffwechselprodukte (Sauerstoff + Zucker) von den Blättern in alle anderen Organe.
Die Wurzeln von Samenpflanzen bilden eine wichtige Struktur, da sie der Wasser– und Nährstoffaufnahme von Pflanzen dienen. Die Abgrenzung der Wurzel von der Sprossachse lässt sich dort ziehen, wo der Übergang der überirdischen Sprossachse in den unterirdischen Teil der Pflanze stattfindet. Das ist jedoch nur ein grober Ansatz, da es auch Samenpflanzen gibt, die epiphytisch wachsen, wie z.B. einige Orchideenarten. Epiphytisch bedeutet hierbei, dass die Pflanzen auf anderen Pflanzen wachsen, ohne direkten Kontakt zum Boden zu haben.
Ein weiteres Unterscheidungsmerkmal von Wurzeln zur Sprossachse liegt darin, dass Wurzeln keine Blätter tragen. In Schwerkraftsexperimenten lässt sich auch ein weiteres Unterscheidungsmerkmal feststellen – die Gravitropie. Gravitropie ist die Eigenschaft von Pflanzen, entweder mit der Schwerkraft oder ihr entgegengesetzt zu wachsen. Wurzeln sind positiv gravitrop, was bedeutet, dass sie in Richtung der Schwerkraft wachsen, während die Sprossachse negativ gravitrop ist, da die Sprossachse entgegen der Richtung der Schwerkraft wächst.

Auch bei umgestürzten Pflanzen wirkt die Gravitropie von Wurzel und Sprossachse
Bei dikotylen Pflanzen herrscht im Wurzelsystem Allorhizie, das bedeutet, dass es eine senkrecht nach unten wachsende Wurzel gibt, aus der seitlich kleinere Wurzeln inklusive Wurzelhaaren abzweigen. Dennoch lässt sich bei Bäumen und Sträuchern feststellen, dass es Arten gibt, bei der eine starke Hauptwurzel vorhanden ist, während andere Arten ein eher oberflächliches Wurzelsystem ausbilden. Diese werden auch als Pfahl- bzw. Flachwurzler bezeichnet.
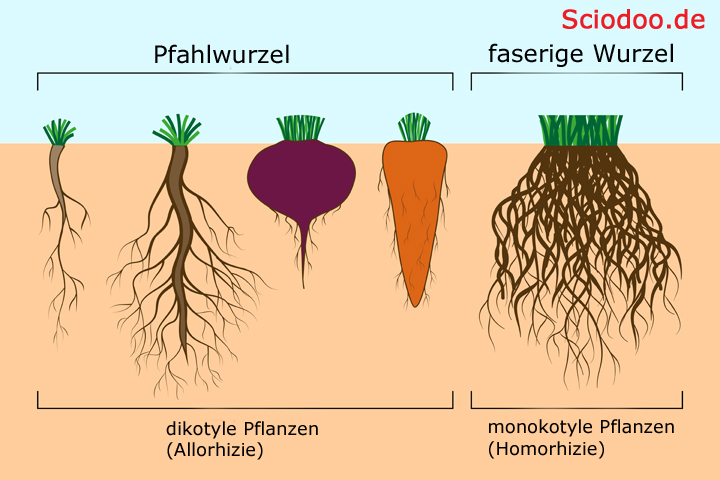
Wurzelsysteme: bei dikotylen Pflanzen (Allorhizie) und monokotylen Pflanzen (Homorhizie)
Beispiele für ein Pfahlwurzelsystem sind u.a. Eichen, Kiefern und Tannen. Pappeln sind als Beispiel für das Flachwurzelsystem heranzuziehen.
Bei monokotylen Pflanzen herrscht Homorhizie, bei der alle Wurzeln gleich stark in alle Richtungen austreiben.
Da die Wurzeln nicht nur der Wasser- und Nährstoffaufnahme dienen, sondern auch eine stabilisierende Funktion für Pflanze und Boden aufweisen, werden Flachwurzler insbesondere in stürmischen Zeiten entwurzelt. Auch bei länger anhaltender Trockenheit haben diese Pflanzen oftmals mit Wassermangel zu kämpfen, da ihre Wurzeln nicht weit genug in den Boden reichen, um an das dort gesammelte Grundwasser zu gelangen.
Wasser- und Nährstoffaufnahme der Wurzeln
Die genauere Funktion der Wasser- und Nährstoffaufnahme lässt sich besser verstehen, wenn man einen Blick auf die Anatomie der Wurzel wirft.
Wurzelspitzenmeristem
Der Ursprung des Wurzelwachstums liegt in einem Bildungsgewebe, dem Wurzelspitzenmeristem. Dieses Wurzelspitzenmeristem liegt geschützt unter einer Haube, der Kalyptra, an der Spitze einer jeden Wurzel und dringt weiter in den Boden ein.

Das Bildungsgewebe (Meristem) in der Wurzelspitze
Damit das Eindringen in den Boden leichter von statten geht, sondert die Kalyptra einen Schleim ab. Dieser Schleim verändert darüber hinaus auch die Bodenstruktur, weshalb gewisse Nährstoffe leichter aufgenommen werden können, andere Schadstoffe jedoch nicht aufgenommen werden. Vom Meristem in Richtung der Wurzel werden die Wurzelzellen vollständig ausgebildet und erlangen ihre natürliche Größe.
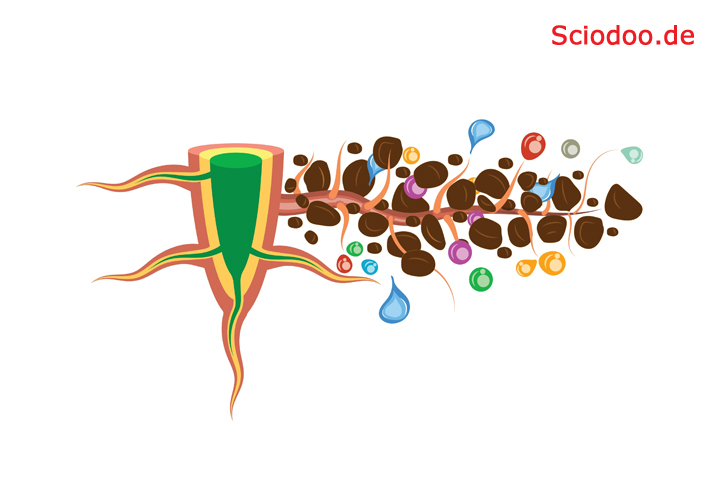
Absorbierung von Nährstoffen und Wasser über die Wurzeln der Pflanzen
Rhizodermis
Die äußere Abgrenzung der Wurzel wird von der Rhizodermis (Wurzelhaut) gebildet. Im Gegensatz zur Oberfläche der Sprossachse fehlen bei der Rhizodermis jedoch die Cuticula (wachsartiger Überzug von Pflanzengeweben wie Spross und Blättern) und die Stomata (Spaltöffnungen).

Zellen an den Wurzelhaaren
Innerhalb der Rhizodermis existiert die Wurzelhaarzone, aus der sogenannte Wurzelhaare gebildet werden können. Die Wurzelhaare haben die Aufgabe, die Wasser- und Nährstoffaufnahme zu optimieren. Das gelingt, der Pflanze unbewusst, durch eine Oberflächenvergrößerung des Wurzelgewebes. Der Zellaufbau der Rhizodermis weist den Charakter eines Speichergewebes auf, da große Vakuolen zu erkennen sind.
Vakuolen sind Zellbestandteile von Pflanzenzellen, die Nährstoffe und Wasser aufnehmen und diese speichern können. Grundsätzlich kann aus jeder Zelle der Rhizodermis ein Wurzelhaar entspringen, dennoch gibt es sogenannte Trichoblasten, die speziell zur Ausbildung von Wurzelhaaren in der Rhizodermis vorliegen. Die Rhizodermis existiert lediglich einige Tage, bis sie letztendlich abstirbt. Abgelöst wird sie von der sogenannten Exodermis.
Exodermis
Die Exodermis wird bereits gebildet, bevor die Rhizodermis abstirbt. Sie besteht aus mehreren Lagen an Zellen, die direkt miteinander verbunden sind. Innerhalb der Zellen wird ein Biopolymer eingelagert, das hydrophobe (wasserabweisende) Eigenschaften aufweist. Dieses Polymer wird als Suberin bezeichnet. Das Suberin verhindert einen unkontrollierten Wasserdurchtritt in die Wurzel und sorgt somit für einen regelbaren Wasserzufluss über den Symplasten.
Wurzelrinde
Die Wurzelrinde umgibt die Exodermis und dient hauptsächlich der Nährstoffaufnahme. Die optimierte Nährstoffversorgung der Pflanze geschieht hierbei bei vielen Pflanzenarten durch diverse Mykorrhiza-Pilze. Mykorrhiza beschreibt eine Symbiose von Pilzen und Pflanzen. Hierbei schließen die Pilze Stickstoff– und Phosphat-Quellen auf, im Gegenzug erhalten sie von der Pflanze Kohlenstoffquellen in Form von Glucose und anderen Zuckern. Die Stickstoffquelle, die von den Pilzen aufgeschlossen wird, ist meist eine Nitratverbindung.
Endodermis
Die Endodermis umschließt innerhalb der Wurzel den Zentralzylinder und stellt die innerste Schicht der Wurzelrinde dar. Sie trägt ebenfalls dazu bei, den geregelten Wasser- und Nährstoffeinstrom in die Wurzel aufrechtzuerhalten, damit dieser nur über den Symplasten erfolgt.
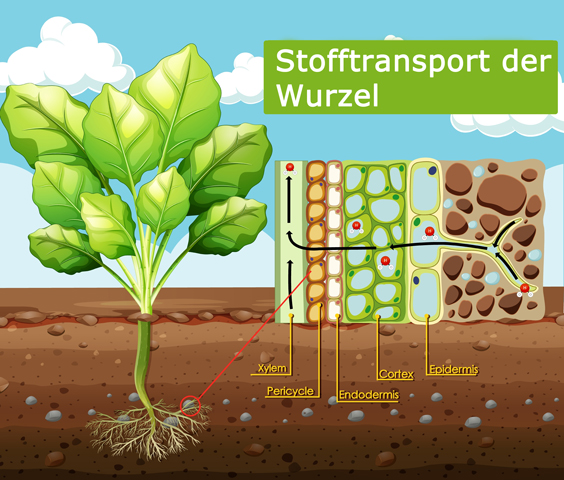
Querschnitt der Wurzel, von rechts nach links. Nährstoffe werden aus dem Erdreich aufgenommen, durchdringen die Epidermis bzw. Exodermis, dann die Rinde (Cortex), die Endodermis, den Perizyklus und gelangen so in die Leitbündel (Xylem). Der Weg über die Plasmodesmen-Verbindungen wird als Symplast bezeichnet
Zentralzylinder und Leitbündel
Der Zentralzylinder, der das innerste der Wurzel darstellt, beinhaltet die Leitbündel. Anhand des Beispiels einer Karotte lässt sich gut verdeutlichen, wo sich der Zentralzylinder befindet. Schneidet man eine Karotte in der Mitte durch und betrachtet die Schnittfläche, so lässt sich eine kreisrunde Struktur im inneren feststellen. Dies ist die Abgrenzung vom Zentralzylinder zur Wurzelrinde.

Querschnitt einer Karotte mit runden Wurzelzylinder als Abgrenzung zum umliegenden Gewebe
Die Leitbündel im Zentralzylinder werden als Xylem und Phloem bezeichnet. Das Xylem transportiert prinzipiell Nährstoffe und Wasser in die anderen Organe der Pflanze, während das Phloem die durch die Photosynthese und andere Reaktionen entstandenen Stoffwechselprodukte aus den Blättern in andere Organe der Pflanze verteilt. Das Xylem ist darüber hinaus aus abgestorbenen, verholzten Zellen aufgebaut, wohingegen das Phloem aus lebenden Zellen besteht, die durch siebartige Strukturen miteinander verbunden sind.
Xylem ist das Leitbündelsystem, welches Wasser und darin gelöste Nährstoffe von der Wurzel in die Pflanze verteilt. Das Phloem, als Leitungssystem, verteilt Stoffwechselprodukte (Sauerstoff + Zucker) von den Blättern in alle anderen Organe.
Pflanzen haben mehrere Möglichkeiten, Nährstoffe und Wasser aus dem Boden aufzunehmen. Drei davon werden im Folgenden näher erläutert:
- Sofern Nährstoffe bereits gelöst im Boden vorliegen, können diese ungehindert vom Symplasten aufgenommen werden.
- Sollten Nährstoffe jedoch an Bestandteile des Bodens gebunden sein, besteht die Möglichkeit einer Austauschabsorption. Hierbei gibt die Pflanze über die Wurzeln Wasserstoffionen und „Kohlensäure“ (CO2 aus der Atmungskette gelöst in Wasser) in den Boden ab und kann somit die gebundenen Moleküle lösen. Nachdem die Moleküle gelöst wurden, stehen diese wie im ersten Fall frei zur Verfügung und können vom Symplasten aufgenommen werden.
- Bei etwas fester gebunden Molekülen, die beispielsweise in Mineralen vorliegen, hilft die Austauschabsorption nicht. Hier muss die Pflanze organische Säuren und Chelat-Bildner (Moleküle, die an mehreren Stellen Metallionen binden) sezernieren (absondern), um diese zu lösen. Im Falle von Pflanzen sind typische Chelatbildner Apfel- bzw. Zitronensäure. Diese umschließen Metallionen und können somit der Pflanze zur Verfügung stehen.
Die Sprossachse
Die Sprossachse dient der Verbindung von Wurzeln und Blättern. Sie ist darüber hinaus dazu in der Lage, die Blätter je nach Sonnenstand so zu bewegen, dass diese immer optimalen Lichtverhältnissen ausgesetzt sind. Weitere Funktionen der Sprossachse sind sowohl die Stabilisierung der Pflanze als auch der Transport und die Speicherung von Wasser und Nährstoffen.
Weitere Bezeichnungen der Sprossachse sind gattungsspezifisch und lauten Halm (bei Gräsern), Stängel (bei kleineren Pflanzen) oder Stamm (bei Sträuchern und Bäumen).
Der Aufbau der Sprossachse ist durchaus vielfältig und je nach Pflanzenfamilie deutlich unterschiedlich. Grundlegend lässt sich die Sprossachse jedoch in das Hypokotyl und das Epikotyl gliedern. Die Kotyledonen (Keimblätter) sind jene Blätter, die nach der Keimung der Pflanze zuerst auftreten. Bei Dikotylen gibt es davon zwei, bei Monokotylen nur eins.
Der Abschnitt zwischen dem Wurzelansatz und dem Keimblatt wird Hypokotyl (Hypo=unterhalb, Kotyl=Keimblatt) genannt. Oberhalb des Keimblattes wird die Sprossachse Epikotyl genannt. Da jedoch die Keimblätter relativ rasch, nach der Ausbildung weiterer Blätter, absterben, kann daraufhin die gesamte Sprossachse als Epikotyl bezeichnet werden.

Während der Keimung unterscheidet man zwei Abschnitte auf der Sprossachse: Hypokotyl und Epikotyl
Eine weitere Begrifflichkeit, die bei der Beschreibung von Pflanzen unerlässlich ist, ist der Nodus bzw. der Internodienabstand. Im Bereich, wo Blätter entspringen, ist die Sprossachse meist verdickt. Diese Verdickung wird aufgrund ihres Aussehens als Nodus bezeichnet (Nodus = Knoten). Der Abstand zwischen zwei Nodi (Plural von Nodus) wird auch Internodienabstand/Internodium genannt.
Die Grundorgane eine Samenpflanze sind Blatt, Wurzel und Sprossachse. Die Blüte ist ein weiteres Organ von Blütenpflanzen.
Diese Abstände spielen insbesondere bei der Beschreibung und Bestimmung von Pflanzen eine essenzielle Rolle. Bei jungen Pflanzen sind die Internodienabstände jedoch nicht exakt zu messen, da diese noch mit dem Pflanzenwachstum größer werden. Mittels interkalaren Wachstums wird die Sprossachse gestreckt.
Weiterhin können auch zwei Arten von Trieben unterschieden werden, sofern beide an einer Pflanze vorhanden sind. Diese werden Lang- und Kurztriebe genannt. Ein Langtrieb besitzt vollständig gestreckte Internodien, während ein Kurztrieb kürzere und gestauchte Internodien aufweist. Beispiele für das Zusammenspiel von Lang- und Kurztrieben lässt sich anhand von Nadelbäumen wie z.B. Kiefern beobachten. Hierbei sind die Langtriebe jene Triebe, die die Kurztriebe tragen, die Nadeln aufweisen.
Verzweigungen der Sprossachse lassen sich bei Samenpflanzen in zwei unterschiedliche Typen gliedern. Diese sind die monopodiale und die sympodiale Verzweigung. Es gibt jedoch einige Ausnahmen, insbesondere sukkulente Pflanzen wie z.B. Kakteen, die kaum Verzweigungen aufweisen.
Die monopodiale Verzweigung stellt eine Art der Verzweigung dar, bei der eine durchgehende Sprossachse vorhanden ist. Hierbei wird jedes Jahr das selbe Spitzenmeristem (Bildungsgewebe an der Spitze der Sprossachse) gefördert, während das Austreiben von Seitentrieben ausbleibt. Ein Beispiel für solche monopodialen Verzweigungen stellt die Fichte dar. Dass hierbei keine Seitentriebe ausgebildet werden, scheint auf den ersten Blick eventuell etwas widersprüchlich, da die Fichte viele Verzweigungen ausbildet. Der Unterschied wird jedoch sichtbar, wenn man die sympodiale Verzweigung als Vergleich heranzieht.

monopodiale Verzweigung einer Sprossachse
Bei der sympodialen Verzweigung wird das Wachstum der abzweigenden Sprosse nicht durch die Hauptachse gefördert, sondern von eigenständigen Seitenachsen. Bei Laubbäumen wie z.B. der Buche lässt sich diese Art der Verzweigung finden. Die endständige Knospe, die sich an der Spitze der Hauptachse befindet, stirbt hierbei ab und es erfolgt das weitere Wachstum aus Seitenknospen.
Werden zwei Seitenachsen gebildet, spricht man von einem Dichasium. Ein solches Dichasium lässt sich beispielsweise beim Flieder finden. Wird hingegen nur eine Seitenachse gebildet, nennt man dies Monochasium. Da diese nahezu in die gleiche Richtung wächst wie die Hauptachse, kann man ein Monochasium auch mit einer monopodialen Verzweigung verwechseln.
Man bezeichnet die scheinbar monopodial verzweigte Achse deshalb auch als Scheinachse. Der Unterschied zwischen Monochasium und einer monopodialen Verzweigung lässt sich an der Anordnung der Blätter festmachen. Seitenachsen enstpringen immer aus der Achsel eines Blattes. Bei einem Monochasium stehen die Blätter an der Scheinachse jedoch den Blütenständen gegenüber. Diese würden jedoch bei einer durchgehenden Hauptachse in den Blattachseln zu finden sein.
Weiterhin kann man zwischen zwei Wuchsformen unterscheiden. Der akrotone Wuchs ist jener, bei dem lediglich die Knospen aus der oberen Sprossregion austreiben. Diese Wuchsform führt zu einer baumtypischen Optik. Der basitone Wuchs hingegen führt zu einer buschähnlichen Optik, da hierbei die Knospen der unteren Sprossregionen zu Seitentrieben ausgebildet werden.
Neben der Charakterisierung der Verzweigungen lassen sich auch unterschiedliche Gewebetypen innerhalb der Sprossachse feststellen. Diese sind das Abschlussgewebe, das Festigungsgewebe, das Grundgewebe und das Leitgewebe. Das Abschlussgewebe enthält unter anderem die Epidermis. Diese kann, ähnlich wie die Epidermis des Blattes, Spaltöffnungen (Stomata) und eine Cuticula besitzen.
Die Zellschicht, die sich unter der Epidermis befindet, bildet die primäre Rinde. Bei Pflanzen, die ein sekundäres Dickenwachstum aufweisen, wird diese primäre Rinde jedoch sehr schnell durch die Borkenschicht ersetzt. Dieser Prozess geht auf die Unfähigkeit der primären Rinde zurück, einem Dickenwachstum folgen zu können. Innerhalb der Borke erfolgt der Gasaustausch, anders als in der Epidermis, über sogenannte Lenticellen.
Das Festigungsgewebe besitzt aufgrund seiner Funktion längliche Zellen mit markanten Zellwänden. Hierbei wird zwischen Sklerenchym- und Kollenchymzellen unterschieden. Das Sklerenchym umgibt die Leitbündel und besteht aus abgestorbenen Zellen. Die oftmals verdickten Zellwände des Sklerenchyms beinhalten oft Lignin, was zu einer Verstärkung des Zellgerüsts führt. Das Kollenchym hingegen ist ein Gewebe aus lebenden Zellen, das nicht verholzt ist. Die Verstärkung dieser Zellen erfolgt durch Polysaccharide (Mehrfachzucker) wie Cellulose oder Pektin.
Das Grundgewebe wird insbesondere durch das Parenchym und dem Mark inmitten des Sprosses gekennzeichnet. Das Parenchym ist hierbei ein organspezifisches Gewebe, das gewisse Aufgaben erfüllt. Es kommt auch in Blättern vor, wo es andere Aufgaben erfüllt als in der Sprossachse. Das Mark der Sprossachse dient vorwiegend der Speicherung von Nährstoffen. Bei einigen Arten zerreißt dieses Mark jedoch, wodurch eine sogenannte Markhöhle entsteht.
Der Transport von Wasser und Nährstoffen geschieht innerhalb des Leitgewebes. Das Xylem und das Phloem sind Bestandteile des Leitgewebes und vollführen unterschiedliche Aufgaben. Im Xylem, das verholzt und aus abgestorbenen Zellen besteht, werden Nährstoffe und Wasser von den Wurzeln zu anderen Organen transportiert.
Innerhalb des Phloems, das aus lebenden Zellen besteht, werden Assimilate (Stoffwechselprodukte) aus der Photosynthese und anderen Reaktionen von den Blättern in andere Organe der Pflanze transportiert. Die Bestandteile des Xylems werden Tracheen und Tracheiden genannt, während man die Bestandteile des Phloems als Siebzellen, Siebröhren und Geleitzellen bezeichnet.
Dennoch können die Leitbündeltypen unterschiedlich gestaltet sein. Es existieren beispielsweise einfache Leitbündel, die nur aus einem Sieb- oder Holzteil bestehen. Darüber hinaus weisen einige Arten zusammengesetzte Leitbündel auf, die einen Sieb- und einen Holzteil besitzen. Die zusammengesetzten Leitbündel lassen sich wiederum in konzentrische und offene Leitbündel unterteilen.
Bei konzentrischen Leitbündeln liegt entweder der Siebteil um den Holzteil oder aber der Holzteil um den Siebteil. Bei dikotylen Pflanzen existieren die offenen Leitbündel, bei denen ein Kambium zwischen dem Xylem und Phloem ausgebildet wird. Das Kambium hat hierbei die Aufgabe, das sekundäre Dickenwachstum zu ermöglichen, ohne dass die Nährstoffversorgung abbricht.
Das Blatt
Das Blatt ist ebenso ein überlebenswichtiges Organ für die Pflanze wie die Wurzeln oder die Sprossachse. Blätter gibt es jedoch nur bei farnartigen Pflanzen und Samenpflanzen. Bei Moosen und Algen werden zwar Gewebe gebildet, die einem Blatt ähneln, aber nicht der Definition eines Blattes entsprechen. Im Laufe der Evolution entstanden aus ursprünglichen Blättern weitere blattähnliche Organe, die jedoch nicht zur Photosynthese geeignet sind. Darunter fallen beispielsweise Blütenblätter oder Blattranken, wie man sie von Kletterpflanzen kennt.
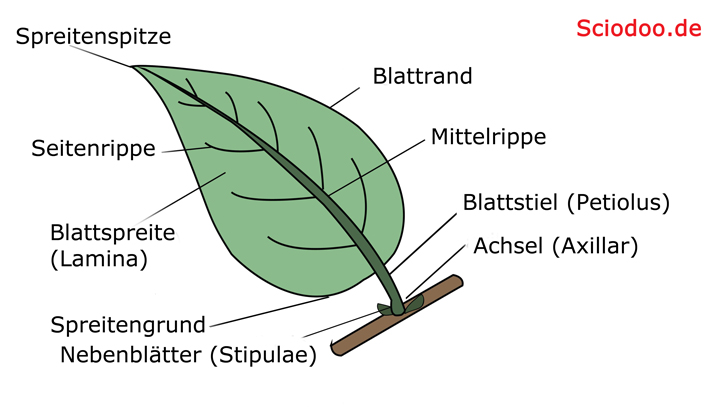
Grundlegende Struktur eines Pflanzenblatts mit Stiel, Spreite und Nebenblättern
Die Anatomie eines Blattes lässt sich in mehrere Gewebe wie z.B. die Epidermis, das Mesophyll und die Leitbündel gliedern. Die Epidermis ist, ähnlich wie bei der Sprossachse, ein Abschlussgewebe. Oftmals ist sie nach außen von einer mehr oder weniger dicken Wachsschicht, der Cuticula, umgeben. Die Cuticula dient dazu, eine unkontrollierte Wasserverdunstung zu verhindern.
Generell, mit einigen Ausnahmen, besitzt die Epidermis keine Chloroplasten und ist somit nicht alleine zur Photosynthese befähigt. Ein Beispiel für solch eine Ausnahme stellen die Schließzellen der Stomata dar, die immer Chloroplasten enthalten, obwohl sie ein Teil der Epidermis sind. Die Stomata sind Spaltöffnungen, die von Schließzellen geöffnet oder geschlossen werden können.
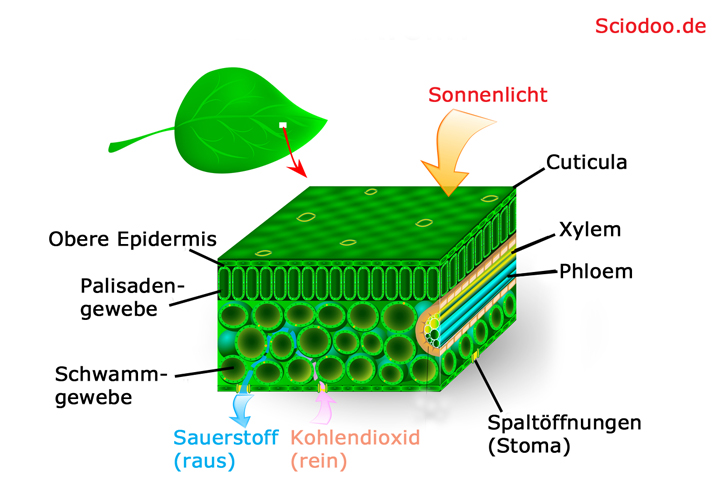
Die verschiedenen Gewebestrukturen in einem Pflanzenblatt, Bei einem Sonnenblatt sind die Cuticula und das Palisadengewebe deutlich dicker
Je nachdem, ob sich die Stomata auf der Ober- oder Unterseite oder auf beiden Seiten des Blattes befinden, nennt man die Blätter epistomatisch (Oberseite, bei Schwimmpflanzen z.B.), hypostomatisch (Unterseite, die häufigste Form) oder amphistomatisch (beide Blattseiten).
Weiterhin können von der Epidermis auch Trichome gebildet werden, das sind haarähnliche Strukturen. Ein Beispiel für solche Trichome weist der Sonnentau auf, der mit seinen klebrigen Drüsenhaaren kleine Insekten fängt. Auch Stacheln, wie sie bei Rosen zu finden sind, sind epidermale Anhänge, die jedoch auch von subepidermalen Zellschichten (Zellschichten, die unterhalb der Epidermis liegen) gebildet werden. In diesem Fall spricht man von Emergenzen statt Trichomen.
Das Mesophyll, auch als Blattparenchym bezeichnet, ist das grundlegende Assimilationsgewebe eines Blattes. Es wird unterteilt in das Palisadenparenchym (Palisadengewebe) und das Schwammparenchym (Schwammgewebe). Das Palisadenparenchym befindet sich in der Regel direkt unter der Epidermis und besitzt aufgrund der Vielzahl an Chloroplasten die Aufgabe, Photosynthese zu betreiben. Das Schwammparenchym dient hauptsächlich der Durchlüftung des Blattgewebes und ist weniger an der Photosynthese beteiligt.
Die Leitbündel sind den Leitbündeln der Sprossachse sehr ähnlich. Auch hier existieren das Xylem und das Phloem. Da die Leitbündel direkt von der Sprossachse in das Blatt übergehen, liegt das Xylem auf der Blattoberseite, während das Phloem auf der Blattunterseite zu finden ist. Die Funktion der Leitbündel im Blatt ist gleich der Funktion der Leitbündel in der Sprossachse – die Wasser- und Nährstoffversorgung.
Die Blüte
Als Blüte bezeichnet man einen kurzen Spross, der unverzweigt vorliegt und nur ein begrenztes Wachstum zeigt. Ein weiteres Merkmal einer Blüte ist die Beteiligung an der sexuellen Fortpflanzung der Pflanze, sei sie direkt oder nur indirekt. Gliedern lässt sich eine Blüte in den Blütenboden, die Kelchblätter, die Kronblätter, die Staubblätter und die Fruchtblätter.
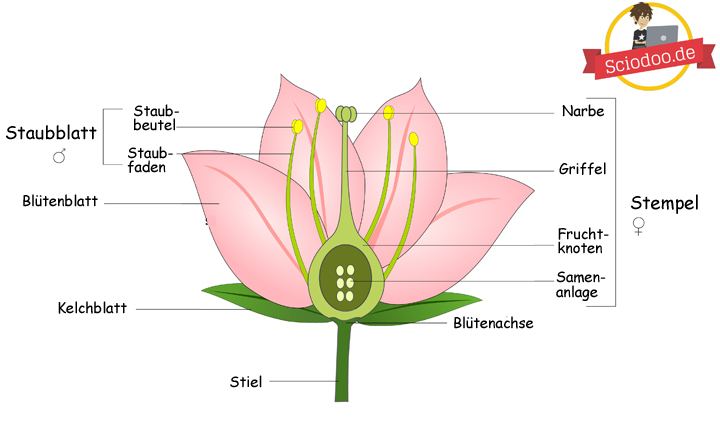
,Grundlegender Aufbau von Blüten einer Blütenpflanze
Die Kelch- und Kronenblätter bilden die Blütenhülle. Diese Blätter sind steril, was bedeutet, dass sie nicht zur Fortpflanzung fähig sind. Solch eine Blütenhülle existiert bei den Angiospermen, jedoch nicht bei den Gymnospermen. Die meistens grün erscheinenden Blätter sind die Kelchblätter, während die Kronblätter oftmals auffällige Farben aufweisen.
Staubblätter sind die Blätter, die die männlichen Fortpflanzungsorgane tragen. Die Fruchtblätter beinhalten hingegen die weiblichen Fortpflanzungsorgane. Nach erfolgter Bestäubung bilden sich in den Fruchtblättern die Samen aus, die letztendlich heranreifen und auf verschiedenste Weisen in der Umwelt verbreitet werden.
Literatur
- Bernhard Kaussmann (Autor), Ulrich Schiewer (Autor), Funktionelle Morphologie und Anatomie der Pflanzen, ISBN: 978-3437204128*
- Julius Wiesner (Autor), Karl Linsbauer (Autor), Anatomie und Physiologie der Pflanzen, ISBN: 978-3368414634*
- Elisabeth Stahl-Biskup (Autor), Jürgen Reichling (Autor), Anatomie und Histologie der Samenpflanzen: Mikroskopisches Praktikum für Pharmazeuten (Wissen und Praxis), ISBN: 978-3769261189*
Über den Autor
